

Mikrofilamentsegenskaper, struktur, funktioner, patologi
- 1285
- 61
- Johan Olsson
De mikrofilament o Actin -filament, är en av de tre grundläggande komponenterna i eukaryota cellcytoskelet.
I eukaryoter är generna som kodar för aktinmikrofilament mycket bevarade i alla organismer, så de används ofta som molekylära markörer för olika studier.
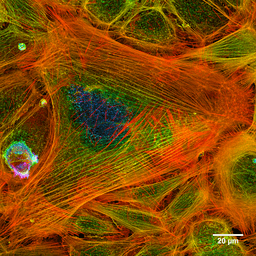
Fotografi av aktinfilamenten av en färgad cell (källa: Howard Vindin [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons) Mikrofilament är fördelade över hela cytosolen, men de är särskilt rikliga i det underliggande området till plasmamembranet, där de bildar ett komplext nätverk och är associerade med andra speciella proteiner för att bilda cytoskeletten.
Mikrofilamentnätverk i cytoplasma av däggdjursceller kodas av två av de sex generna som beskrivs för aktin, som är involverade i dynamiken i mikrofilament och som till och med är mycket viktiga under differentieringen av stamcellerna.
Många författare är överens om att mikrofilament är de mest mångsidiga, mångsidiga och viktiga proteiner från cytoskeletten i de flesta eukaryota celler, och det är viktigt att komma ihåg att dessa inte finns i prokaryota mikroorganismer.
I denna typ av celler finns det emellertid homologa filament till mikrofilament, men som bildas av ett annat protein: MREB -protein.
För närvarande tros det att genen som kodar för detta protein är en möjlig förfädergen för eukaryota aktin. Emellertid är aminosyrasekvenshomologin som bildar MREB -protein endast 15% med avseende på aktinsekvensen.
Liksom är en grundläggande del av cytoskeletten, kan alla fenotypiska defekt både i mikrotubuli och i mellanliggande filament och aktinmikrofilament (cytoskelett) orsaka olika cell- och systempatologier.
[TOC]
Egenskaper och struktur
Mikrofilament består av proteinmonomerer i aktinfamiljen, som är extremt rikliga kontraktila proteiner i eukaryota celler, eftersom de också deltar i muskelkontraktion.
Dessa filament har en diameter mellan 5 och 7 nm, så de är också kända som tunna filament och består av två former av aktin: den globulära formen (aktin G) och den filamentösa formen (aktin F).
Proteiner som deltar i cytoskeletten är kända som y- och ß -aktiner, medan de som deltar i sammandragning vanligtvis är aktiner a.
Kan tjäna dig: cytoplasma: funktioner, delar och egenskaperAndelen globulära aktin och filamentösa aktin i cytosolen beror på cellulära behov, eftersom mikrofilament är mycket varierande och mångsidiga strukturer, som ständigt tillväxt och förkortas på grund av polymerisation och depolymeterisering.
Actin G är ett litet kulaprotein, bestående av nästan 400 aminosyror och cirka 43 kDa molekylvikt.
Actin g monomerer som utgör mikrofilamenten beställs i form av en spiralformad sträng, eftersom var och en lider av en vridning när de är associerade med följande.
Aktin G är associerad med en Ca2+ -molekyl och en annan ATP, som stabiliserar dess kulformiga form; Medan aktin F erhålls efter hydrolysen av det terminala fosfatet i ATP -molekylen i aktin G, vilket bidrar till polymerisation.
Organisation
Actin -filament kan organiseras i form av "strålar" eller "nätverk" som har olika funktioner i cellerna. Strålarna bildar parallella strukturer kopplade av ganska styva tvärbroar.
Nätverk är å andra sidan mer bekväma strukturer, till exempel tre dimensionella nät med egenskaperna hos semi -solida geler.
Det finns många proteiner som är associerade med aktin- eller mikrofilamentfilament och som är kända som ABP (från engelska Aktinbindande proteiner), som har specifika webbplatser för detta.
Många av dessa proteiner tillåter mikrofilament att interagera med de andra två komponenterna i cytoskeletten: mikrotubulor och mellanliggande filament, liksom med de andra komponenterna i plasmamembranets inre yta.
Bland andra proteiner som interagerar med mikrofilament är kärnark och spektrin (i röda blodkroppar).
Hur bildas aktinfilament?
Eftersom globulära aktinmonomerer alltid går med på samma sätt, orienterade i samma riktning, har mikrofilament en definierad polaritet, med två ytterligheter: en "mer" och en "mindre".
Polariteten hos dessa filament är mycket viktig, eftersom de växer betydligt snabbare på grund av deras positiva slut, där de nya aktinerna monomerer läggs till.
 Grafisk representation av aktinmikrofilament (källa: derivatarbete: retama (samtal) thin_filament_formation.SVG: Mikael Hägström [Public Domain] via Wikimedia Commons)
Grafisk representation av aktinmikrofilament (källa: derivatarbete: retama (samtal) thin_filament_formation.SVG: Mikael Hägström [Public Domain] via Wikimedia Commons) Det första som äger rum under polymerisationen av aktinfilament är en process som kallas "kärnbildning", som består av föreningen av tre proteinmonomerer.
Kan tjäna dig: plasomolysTill denna trimmer läggs nya monomerer till i båda ändarna, så att filamentet växer. Aktin G-monomerer kan hydrolyz ATP med varje förening, vilket har konsekvenser för polymerisationshastigheten, eftersom aktin-ATP-delar dissocierar mer svårigheter än aktin-ADP.
ATP är inte nödvändig för polymerisation och den konkreta funktionen hos dess hydrolys har ännu inte klargjorts.
Vissa författare anser att eftersom aktinpolymerisationshändelser är snabba.
Reglering
Både polymerisationen av aktinfilament och dess depolimerisering är processer som är mycket reglerade av en serie specifika proteiner, som är ansvariga för ombyggnaden av filamenten.
Exempel på proteinerna som reglerar depolymerer. Ett annat protein, profylain, har en motsatt funktion, eftersom det stimulerar föreningen mellan monomerer (genom att stimulera ADP: s utbyte mot ATP).
Funktioner
Mikrofilament interagerar med myosinfilament som är associerade med transmembranproteiner som har en domän i cytosol och en annan utomlandscell ute, så de deltar i cellmobilitetsprocesser.
Dessa mikrofilament associerade med plasmamembranet förmedlar olika cellulära svar på olika typer av stimuli. Till exempel är cell vidhäftning av epitelvävnader riktade av transmembranproteiner kända som cadherinas, som interagerar med mikrofilament för att rekrytera svarfaktorer.
Aktinfilament interagerar med mellanliggande filament för att göra extracellulära stimuli överför till viktiga platser som ribosomer och kromosomer inuti kärnan.
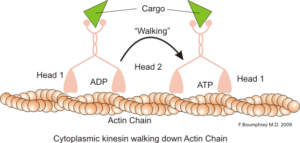 Representation av den intracellulära motoriska funktionen hos aktinmikrofilament (källa: BOUMPHREYFR [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)] via Wikimedia Commons)
Representation av den intracellulära motoriska funktionen hos aktinmikrofilament (källa: BOUMPHREYFR [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)] via Wikimedia Commons) En klassisk och mycket studerad funktion av mikrofilament är dess förmåga att bilda "broar", "skenor" eller "motorvägar" för rörelse av myosinmotorprotein I, som kan ladda transportvesiklar från organisationerna till membranplasma på det sekretory vägar.
Mikrofilament interagerar också med myosin II för att etablera den kontraktila ringen som bildas under cytokinesis, just under det sista steget av celldelningen där cytosolen separeras från stam- och dottercellerna.
Kan tjäna dig: polymorfonukleära leukocyterI allmänhet modulerar aktinformade mikrofilament för fördelningen av vissa organeller såsom Golgi -komplexet, endoplasmatisk retikulum och mitokondrier. Dessutom deltar de också i den rumsliga positioneringen av RNMS så att de läses av ribosomerna.
Hela mobiltelefonuppsättningen av mikrofilament, särskilt de som är nära besläktade med plasmamembranet, deltar i bildningen av böljande cellmembran av celler som har en konstant aktiv rörelse.
De deltar också i bildandet av mikrovningar och andra vanliga utsprång på ytan på många celler.
Exempel på funktioner i levern
Mikrofilament deltar i gallsekretionsprocessen i hepatocyter (leverceller) och även i peristaltiska rörelser (koordinerad sammandragning) av leverkanalikuli.
De bidrar till differentieringen av plasmamembrandomäner tack vare deras associering med olika cytosoliska element och kontrollen de utövar på topografin i dessa intracellulära element.
Relaterade patologier
Det finns få sjukdomar associerade med primära defekter i strukturen eller med proteiner och reglerande enzymer i syntesen av mikrofilament, även om dessa är direkt involverade i ett stort antal funktioner.
Det låga indexet för sjukdomar och missbildningar i den primära strukturen för mikrofilament beror på att det i allmänhet finns flera kodande gener av både aktin och dess reglerande proteiner, ett fenomen som kallas "genetisk redundans".
En av de mest studerade patologin är förglasning av oocyter på deras cytoskelett, där ett avbrott observeras i det kortikala mikrofilamentnätverket, liksom en depolimerisering och en desorganisering av mikrotubulorna i den mitotiska spindeln.
I allmänna termer orsakar denna förglasning kromosomal spridning, eftersom det leder till en oenighet i komprimeringen av hela kromatinet.
Cellerna som har en större organisation och en del av mikrofilament i deras cytoskelett är strippade muskelceller, därför är de flesta patologier associerade med en funktionsfel i den kontraktila apparaten.
Defekta eller atypiska mikrofilament har också associerats med bensjukdom känd som Paget -sjukdom.
Referenser
- Aguilar-Cuenca, R., Llorente-González, c., Vicente, c., & Vicente-Manzanares, M. (2017). Mikrofilamentkoordinerad vidhäftningsdynamik driver en cellmigration och former vem vävnader. F1000Resarch, 6.
- Två åtgärder, c. G., Chhabra, D., Kekic, m., Finger, jag. V., Tsubakihara, m., Bär, D. TILL., & Nosworthy, n. J. (2003). Bindande proteiner aktin: Reglering av cytoskeletala mikrofilament. Fysiologiska recensioner, 83(2), 433-473.
- Guo, h., Fauci, l., Shelley, m., & Kanso, och. (2018). Bistabilitet vid synkronisering av aktiverade mikrofilament. Journal of Fluid Mechanics, 836, 304-323.
- Lansera., Langer, r., & Vacanti, J. P. (Eds.). (2011). Principer för vävnadsteknik. Akademisk press.
- Robbins, j. (2017). Sjukdomar i cytoskeletten: deminopatierna. I hjärtskeletala myopatier hos barn och du är vuxna (pp. 173-192). Akademisk press.
- « Grizzly Bear -egenskaper, livsmiljö, mat, beteende
- Cistus Clusii -egenskaper, taxonomi, livsmiljö, distribution, användningar »