

Purinernas egenskaper, struktur, funktioner
- 2238
- 100
- Prof. Erik Johansson
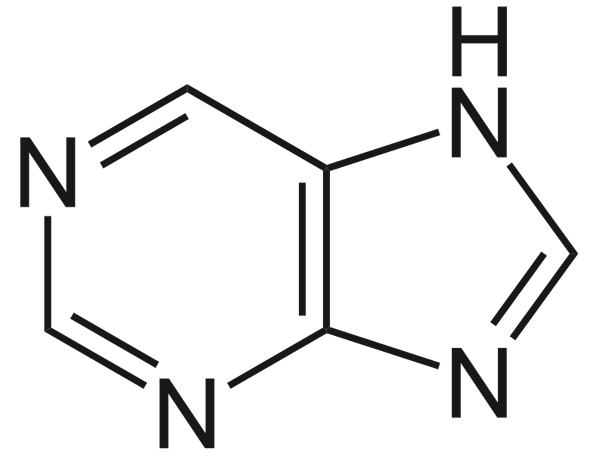
De Purinor De är strukturellt platta, heterocykliska molekyler, bildade av sammansmältningen av två ringar: en av sex atomer och en annan av fem. De viktigaste molekylerna som inkluderar puriner är nukleotider. De senare är blocken som ingår i nukleinsyror.
Förutom deras deltagande i arvsmolekylerna finns puriner närvarande i högenergi -strukturer såsom ATP och GTP och andra molekyler av biologiskt intresse, såsom nikotinamidadenin -dinukleotid, nikotinamidadenin dinukleotidfosfat (NADPH) och koenzym Q Q.
Källa: Sponk [Public Domain] [TOC]
Egenskaper och struktur
Strukturen av puriner är som följer: en heterocyklisk molekyl, bildad av en pyrimidinring och en imidazolring. När det gäller antalet atomer har ringarna sex och fem atomer.
De är platta molekyler som innehåller kväve. Vi hittar dem som en del av nukleosider och nukleotider. De senare är de strukturella blocken av nukleinsyror: DNA och RNA.
Hos däggdjur är puriner i större andel i DNA- och RNA -molekyler, särskilt som adenin och guanina. Vi hittar dem också i unika molekyler som AMP, ADP, ATP och GTP, bland andra.
Funktioner
-Strukturella block av nukleinsyror
Nukleinsyror ansvarar för att lagra genetisk information och orkestrera proteinsyntesprocessen. Strukturellt sett är de biopolymerer vars monomerer är nukleotider.
Purinas är en del av nukleotider
I en nukleotid hittar vi tre komponenter: (1) en fosfatgrupp, (2) ett fem -kolsocker och (3) en kvävebas; Socker är den centrala komponenten i molekylen.
Kvävebasen kan vara en purin eller en pyrimidin. Purinerna som vi normalt finner i nukleinsyror är guanin och adenin. Båda är ringar sammansatta av nio atomer.
Puriner bildar glukosida kopplingar med ribos genom kväve i position 9 och kol 1 av socker.
En anglo -Saxon nemonisk regel att komma ihåg att puriner har nio atomer är att båda termerna på engelska, Adenin och Guanin De har ordet Nio, Vilket betyder nio.
Det kan tjäna dig: Histologi: Historia, vilka studie- och studiemetoderPurinas parar sig inte med varandra
Den dubbla DNA -propellen kräver parning av baserna. På grund av ett steriskt hinder (det vill säga av storleksskäl) kan en purin inte vara blandad av en annan purin.
Under normala förhållanden framgår adenin purin med tyminpyrimidin (A + T) och guaninpurin med cytosinpyrimidin (G + C). Kom ihåg att pyrimidiner är platta molekyler som består av en enda ring och därför mindre. Detta mönster kallas Chargoffs regel.
Strukturen för RNA -molekylen består inte av en dubbel propell, men ändå hittar vi samma puriner som vi nämner i DNA. Kvävebaserna som varierar mellan båda molekylerna är pyrimidiner.
-Energilagringsmolekyler
Tre -fas nukleosider, särskilt ATP. De allra flesta kemiska reaktioner i metabolism använder energin lagrad i ATP.
Bindningarna mellan fosfater är hög energi, Eftersom flera negativa avgifter tillsammans avvisar och gynnar uppdelningen av samma. Energin som släpps används av cellen.
Förutom ATP är puriner beståndsdelar av biologiska intressemolekyler såsom nikotinamidadenin dinukleotid, nikotinamid adenin dinukleotidfosfat (NADPH) och koenzym Q.
-Neurotransmittorer
Många studier har visat att puriner fungerar som signalmolekyler för Glia i centrala nervsystemet.
Puriner kan också hittas som en del av strukturer som kallas nukleosider. De liknar nu nukleotider, men saknar fosfatgruppen.
Nukleosider har lite relevant biologisk aktivitet. Men hos däggdjur finner vi ett mycket markant undantag: adenosin. Denna molekyl har flera funktioner och är involverad i regleringen av processer i det nervösa och kardiovaskulära systemet, bland andra.
Det kan tjäna dig: Flora och Fauna of Santa Fe: Representativa arterAdenosinens handling i sömnreglering är välkänd. I hjärnan hittar vi flera receptorer för denna nukleosid. Närvaron av adenosin är relaterad till känslan av trötthet.
Purinmetabolism
Syntes
Biosyntesen av purinerna initieras med ett ribos-5-fosfatskelett. Fosforbosylenzymsyntetaspyrofosfatet ansvarar för att katalysera tillsatsen av ett pyrofosfat.
Därefter är det glutamin-PRPP-enzymet amidotransferas eller amidofosforribosyltransferas, som katalyserar interaktionen mellan PRPP (förkortning för att utse föreningen som produceras i föregående steg, fosforribosylpyrofosfat) och glutamin för att bilda produkten 5-fosforbosylamine.
Den senare föreningen fungerar som ett skelett för en serie molekylära tillägg, vars sista steg är bildandet av monofosfatinosin, förkortat som omöjligt.
IMP kan fortsätta till omvandlingen av AMP eller GMP. Dessa strukturer kan fosforyleras för skapandet av högenergimolekyl, såsom ATP eller GTP. Denna rutt består av 10 enzymatiska reaktioner.
I allmänhet är hela processen med purinsyntes mycket energiberoende, så det kräver konsumtion av flera ATP -molekyler. Syntes novo av puriner förekommer mestadels i cytoplasma i leverceller.
Dietkrav
Både puriner och pyrimidiner produceras i adekvata mängder i cellen, så det finns inga oundgängliga krav för dessa molekyler i kosten. Men när dessa ämnen konsumeras återvinns de.
Sjukdomar förknippade med purinmetabolism: gikt
Inuti cellen är ett av resultaten av metabolismen av de puriska baserna produktionen av urinsyra (c5H4N4ANTINGEN3), På grund av verkan av ett enzym som kallas xantina oxidas.
Hos en frisk person är det normalt att hitta låga koncentrationer av urinsyra i blodet och urinen. Men när dessa normala värden blir höga, ackumuleras detta ämne gradvis i kroppens leder och i vissa organ, till exempel njurarna.
Det kan tjäna dig: Zygote: Klassificering, utbildning, utveckling och segmenteringDietens sammansättning är en avgörande faktor i produktionen av gikt, eftersom intaget fortsätter av rika purinelement (alkohol, rött kött, marik, fisk, bland andra) kan i sin tur öka urinsyrakoncentrationer.
Symtomen på detta tillstånd är rodnaden i de drabbade områdena och en intensiv smärta. Det är en av de typer av artrit som påverkar patienter genom ackumulering av mikrokristaller.
Referenser
- Alberts, b., Bray, D., Hopkin, K., Johnson, A. D., Lewis, J., Raff, m.,... & Walter, s. (2013). Essential Cell Biology. Kransvetenskap.
- Borea, s. TILL., Gessi, s., Meright, s., Vincenzi, f., & Varani, K. (2018). Farmakologi av adenosinmottagare: The State of the Art. Fysiologiska recensioner, 98(3), 1591-1625.
- Brady, s. (2011). Grundläggande neurokemi: Principer för molekylär, cellulär och medicinsk neurobiologi. Akademisk press.
- Cooper, g. M., & Hausman, r. OCH. (2007). Cellen: En metodmolekylär. Washington, DC, Sunderland, MA.
- Devlin, t. M. (2004). Biokemi: Lärobok med kliniska tillämpningar. Jag reverserade.
- Firesin, g. S., Budd, r., Gabriel, s. OCH., McInnes, jag. B., & O'Dell, J. R. (2016). Kelley och Firesins lärobok om reumatology e-bok. Elsevier Health Sciences.
- Griffiths, a. J. (2002). Modern genetisk analys: integrera gener och genom. Macmillan.
- Griffiths, a. J., Wessler, s. R., Lewontin, r. C., Gelbart, w. M., Suzuki, D. T., & Miller, J. H. (2005). En introduktion till genetisk analys. Macmillan.
- Koolman, J., & Röhm, K. H. (2005). Biokemi: Text och Atlas. Ed. Pan -amerikansk medicin.
- Mikhailopulo, jag. TILL., & Miroshnikov, a. Yo. (2010). Nya trender inom nukleosidbioteknik. Naturae 2 minuter(5).
- Passarge, E. (2009). Genetik text och atlas. Ed. Pan -amerikansk medicin.
- Pelley, J. W. (2007). Elseviers integrerade biokemi. Mosby.
- Siegel, g. J. (1999). Grundläggande neurokemi: Molekylära, cellulära och medicinska aspekter. Lippincott-raven.
- « Cytogenetikhistoria, vilka studier, tekniker, applikationer
- Fosfatidilinositolstruktur, träning, funktioner »