

Muscariniska receptorer struktur, typer och deras funktioner, antagonister
- 3574
- 591
- Hans Olsson
De muskarinreceptorer De är molekyler som förmedlar acetylkolin (ACH) och är belägna i det postsynaptiska membranet av synapser där nämnda neurotransmitter frisätts; Hans namn kommer från hans känslighet för den muskarinalkaloid som produceras av svampen Amanita muscaria.
I det centrala nervsystemet finns det flera neuronala uppsättningar vars axoner frigör acetylkolin. Några av dem slutar i hjärnan själv, medan de flesta utgör motorvägarna för skelettmuskler eller effektorvägarna i det autonoma nervsystemet för körtlarna och hjärtat och släta muskler.
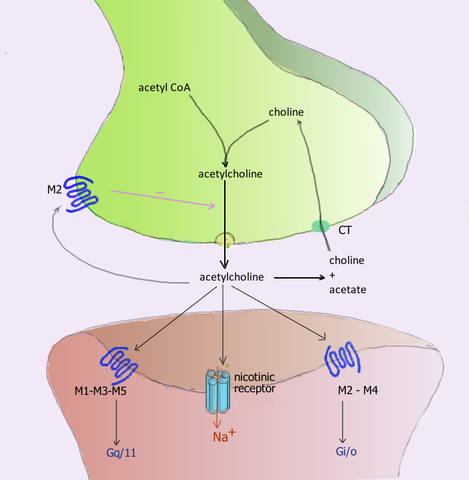
Acetylkolin neuroreceptor under synapsen och deras respektive receptorer i det postsynaptiska membranet (källa: Användare: Pancrat [GFDL 1.2 (http: // www.gnu.Org/licenser/gamla licenser/FDL-1.2.html)] via Wikimedia Commons) Acetylkolin frisatt i de neuromuskulära fackföreningarna i de aktiva skelettmuskelkolinergiska receptorerna som kallas nikotinik, för dess känslighet för nikotinalkaloiden, och som också finns i synaperna ganglioner i det autonoma nervsystemet (SNA).
De postganglioniska neuronerna i den parasympatiska uppdelningen av detta system utövar deras funktioner genom att frigöra acetylkolin, som verkar på de muskariniska kolinergiska receptorerna belägna i membranen i effektorcellerna och inducera i dem elektriska modifieringar genom förändringar av permeabilitet i deras joniska kanel.
 Kemisk struktur för acetylkolin neurotransmitter (källa: Neurotokeker [public domain] via Wikimedia Commons) [TOC]
Kemisk struktur för acetylkolin neurotransmitter (källa: Neurotokeker [public domain] via Wikimedia Commons) [TOC]
Strukturera
De muskariniska receptorerna tillhör familjen till de metabotropa receptorerna, en term med vilka de receptorer som inte är korrekt joniska kanaler betecknas, utan proteinstrukturer som när det är aktiverat trigger intracellulära metaboliska processer som modifierar verksamheten hos riktiga kanaler.
Termen används för att skilja dem från jonotropiska receptorer, som är riktiga jonkanaler som öppnar eller stänger av direktverkan av neurotransmitteren, som är fallet med de nikotiniska receptorerna som redan nämns av de neuromuskulära placken i skelettmuskeln.
Inom de metabotropa receptorerna ingår muskariniker i gruppen som kallas Grozed G de aktiverar fosfolipas C (PLC).
Muskarinreceptorer är långa omfattande membranproteiner; De har sju transmarketssegment som består av alfa -propens, som i följd korsar lipidmembranalskiktet. Inuti, på det cytoplasmiska ansiktet, är de associerade med motsvarande G-protein som överför ligand-mottagarinteraktionen.
Det kan tjäna dig: Embrioblaster: Utveckling, funktioner och lagerTyper av muskarinreceptorer och deras funktioner
Minst 5 typer av muskarinreceptorer som är betecknade med bokstaven M följt av ett antal har identifierats, nämligen: M1, M2, M3, M4 och M5.
M1-, M3- och M5 -receptorer bildar M1 -familjen och kännetecknas av deras GQ- eller G11 -proteinassociation, medan M2- och M4 -receptorer kommer från M2 -familjen och är associerade med GI -proteinet.
- M1 -receptorer
De finns främst i centrala nervsystemet, i de exokrina körtlarna och i nodon autonoma system ganglia. De är kopplade till GQ -protein, som aktiverar fosfolipas C -enzymet, som omvandlas till fosfatidylinositolen (PIP2) till inositol tryposfat (IP3), som frigör intracellulära, och diacilglycerol (DAG), som aktiverar proteiner C C.
- M2 -receptorer
De finns i grunden i hjärtat, främst i cellerna i den kinesiska noden, på vilken de verkar genom att minska sin urladdningsfrekvens, som beskrivs nedan.
Hjärtautomatism
M2 -receptorer har studerats mer grundligt vid hjärtnoden (SA) i hjärtat, där automatiken som regelbundet producerar de rytmiska exciteringarna som är ansvariga för hjärtmekanisk aktivitet normalt manifesteras.
Cellerna i den kinesoaurikulära noden, efter varje verkningspotential (PA) som utlöser en hjärtsystol (sammandragning), ompolariseras och återgår till nivån på cirka -70 mV. Men spänningen förblir inte i det värdet, men lider progressiv depolarisering till en tröskelnivå som utlöser en ny handlingspotential.
Denna progressiva depolarisering beror på spontana förändringar i jonströmmar (i) som inkluderar: K+ (IK1) utgångsreduktion, utseende av en inträdesström på Na+ (FI) och sedan en inträde av Ca ++ (ICAT), tills de når tröskeln och en annan CA ++ (ICAL) ström som ansvarar för handlingspotentialen utlöses.
Om utgången från K+ (IK1) är mycket låg och ingångsströmmarna för Na+ (IF) och Ca ++ (icat) är hög, sker depolarisering snabbare, handlingspotential och sammandragning inträffar tidigare och frekvenshjärtan är högre. Motsatta modifieringar i dessa strömmar lägre frekvens.
Kan tjäna dig: CariocinesisMetabotropa förändringar inducerade av norepinefrin (sympatisk) och acetylkolin (parasympatisk) kan förändra dessa strömmar. AMPC aktiverar direkt IF -kanalerna, proteiner A (PKA) fosforyila och aktiverar ICAT Ca ++ -kanaler och p GI -proteingruppen aktiverar utgången från K+.
Muscarinic Action M2
När acetylkolin frisatt av de postganglioniska termineringarna av hjärtvagala fibrer (parasympatiska) binder till M2 -muskarinreceptorerna för cellerna i den sinoaurikulära noden, ändrar GI -proteinet αi -proteinet, och separerar, lämnar blocket ß -yy.
AI -subenheten hämmar adenylciklas. Detta sista faktum minskar fosforylering och aktivitet av Ca ++ -kanaler för ICAT; Resultatet är en minskning av depolariserande strömmar.
Gruppen som bildas av py -subenheterna i GI -proteinet aktiverar en ström av K+ ut (IKACH) som tenderar att motverka ingångarna till Na+ och Ca ++ och sänka depolarisationshastigheten.
Det gemensamma resultatet är en minskning av lutningen av spontan depolarisering och en minskning av hjärtfrekvensen.
- M3 -receptorer
 M3 Muscarinic Receptor Scheme (Källa: Takuma-SA [CC0] via Wikimedia Commons)
M3 Muscarinic Receptor Scheme (Källa: Takuma-SA [CC0] via Wikimedia Commons) De finns i den glatta muskeln (matsmältningssystem, urinblåsan, blodkärl, bronki), i vissa exokrina körtlar och i centrala nervsystemet.
GQ -protein är också kopplade och på lungnivån kan de orsaka bronkokonstriktion, medan de verkar i det vaskulära endotelfrisättningen kväveoxid (NO) och orsaka vasodilatation.
- M4- och M5 -receptorer
Dessa receptorer är mindre karakteriserade och studerade än de tidigare. Hans närvaro i centrala nervsystemet och i vissa perifera vävnader har rapporterats, men deras funktioner är inte tydligt etablerade.
Kan tjäna dig: Tata Box: Egenskaper och funktionerAntagonister
Den universella antagonisten för dessa receptorer är atropin, en alkaloid extraherad från växten Belladon atropa, som binder till dem med en hög affinitet, som representerar ett kriterium för att skilja dem från nikotinreceptorer som är okänsliga för denna molekyl.
Det finns ett stort antal andra antagonistiska ämnen som binder till de olika typerna av muskarinreceptorer med olika affiniteter. Kombinationen av olika affinitetsvärden för några av dem har tjänat exakt för att inkludera dessa receptorer i en eller annan av de beskrivna kategorierna.
En partiell lista över andra antagonister skulle inkludera: pirenzepin, metotektramin, 4-damp, hymbacin, AF-DX 384, tripitramin, darifenacin, PD 102807, AQ RA 741, PFHHSID, MT3 och MT7; toxiner de senare i gifterna med de gröna respektive svarta mambaserna.
M1 -receptorer har till exempel hög känslighet för pirenzepin; M2 av tripitramin, metktramin och hybacin; M3 med 4-damp; M4 är mycket relaterade till MT3 -toxin och även till hybacin; M5 liknar M3, men med avseende på dem är de mindre relaterade för här 741.
Referenser
- Ganong WF: Neurotransmittorer och neuromodulatorer, i: Granskning av medicinsk fysiologi, 25: e upplagan. New York, McGraw-Hill Education, 2016.
- González JC: Muscariniska receptorer i moduleringen av GABAergisk överföring i hippocampus. Minne för att välja läkarexamen. Autonomous University of Madrid. 2013.
- Guyton AC, Hall JE: Rythmical Excitation of the Heart, In: Lärobok för medicinsk fysiologi , 13: e upplagan; AC Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Piper HM: Herzerregung, i: Physiologie des Menschen Mite Pathophysiologie, 31: e upplagan; RF Schmidt et al (eds). Heidelberg, Springer Medizin Verlag, 2010.
- Schrader J, Gödeche A, Kelm M: Das Hertz, i: Fysiologi, 6: e upplagan; R KLINKE et al (eds). Stuttgart, Georg Thieme Verlag, 2010.
- Siegelbaum SA, Clafam DE, Schwartz JH: Modulering av synaptisk överföring: Second Messengers, In: Principles of Neural Science, 5th ed; E Kandel et al (eds). New York, McGraw-Hill, 2013.
- « Cellularbiologihistoria, vilka studier, applikationer och koncept
- Hur reproducerar växter? Asexuell och sexuell reproduktion »