

Joniska kanaler struktur, funktioner, typer
- 961
- 135
- Lars Eriksson
De jonkanaler Det är ihåliga membranala strukturer som bildar kanaler eller porer som korsar membranets tjocklek och kommunicerar utsidan av cellen med dess cytosol och vice versa; Vissa kan ha ett grindsystem som reglerar dess öppning.
Dessa kanaler är fulla av vatten och kontrollerar passagen av specifika joner från ena sidan till den andra av membranet. De bildas av proteiner som är typiska för cellmembran som bildar cylindriska rörformade strukturer som går igenom dem.
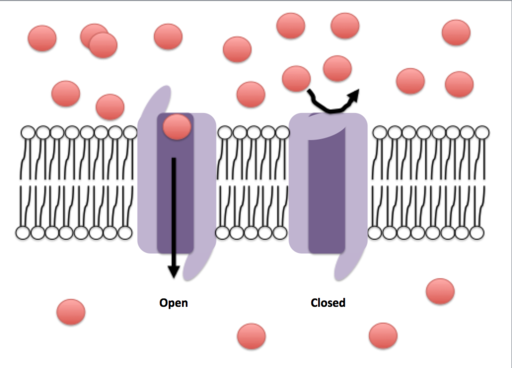
 Öppen och stängd konformation av en jonisk kanal (källa: Efazzari [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons)
Öppen och stängd konformation av en jonisk kanal (källa: Efazzari [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons) Transportmekanismer genom dessa membran kan klassificeras vid passiv eller aktiv transport. Skulderna är de som tillåter passering av ämnen till förmån för deras koncentrationsgradienter, under tiden kräver tillgångarna energiförbrukning, eftersom de förskjuter ämnen mot sina koncentrationsgradienter.
Joniska kanaler utgör en passiv transportmekanism som kan klassificeras enligt deras specificitet, det vill säga enligt vilken typ av jon de låter passera, eller beroende på hur de öppnar eller stänger eller stänger.
https: // giphy.com/gifs/jon-kanal-y07c7ocoigykjl6tzz
Huvudfunktionen för dessa membranala transportsystem är att tillåta den reglerade passagen av ämnen i eller utanför cellerna och därmed upprätthålla intracellulära koncentrationer av joner och andra ämnen.
Närvaron av cellmembran och joniska kanaler i grunden för att upprätthålla koncentrationsskillnader mellan intracellulära och extracellulära medier, som har relevans från många synpunkter.
Joniska kanaler, särskilt de beroende liganden, är mycket viktiga inom farmakologi och medicin, eftersom många läkemedel kan efterlikna funktionerna hos naturliga ligander och gå med i kanalen, öppna eller stänga den, eftersom fallet kan vara fallet.
Andra läkemedel kan blockera unionens plats och därmed förhindra verkan av naturlig ligand.
[TOC]
Strukturera
Strukturen för joniska kanaler bildas av specifika transmembranproteiner som har en tubulär form och lämnar en por eller hål som möjliggör kommunikation mellan interiören och utsidan av cellen eller mellan intracellulära fack (organeller).
Varje jonkanal innebär ett specifikt strukturellt protein i membranet och mer än 100 gener som kodar för specifika jonkanaler har beskrivits.
För natriumkanalen, till exempel 10 gener som heter SCN som kodar olika proteiner fördelade i olika vävnader med särskilda funktioner och strukturer.
Likaså en betydande mängd gener som kodifierar olika proteiner som utgör kaliumkanaler som tillhör olika familjer och har olika mekanismer för aktivering, öppning och inaktivering.
Proteinstruktur för en jonisk kanal
Vanligtvis består en funktionell jonkanal associerad med ett membran av sammansättningen av 4 till 6 liknande polypeptidsubenheter (HOMO -oligomerer) eller olika (hetero -oligomerer) som bildar en central por mellan dem.
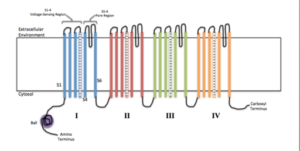 Schema för membranala underenheter av en jonisk kanal (källa: efazzari [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons)
Schema för membranala underenheter av en jonisk kanal (källa: efazzari [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons) Varje underenhet varierar beroende på kanalens egenskaper och egenskaper, eftersom många är specifika för vissa joner och har olika öppnings- och stängningsmekanismer.
Kan tjäna dig: cnidocyter: egenskaper, struktur, funktioner, typerVissa kanaler består av en enda polypeptidkedja organiserad i upprepade motiv som går igenom membrantjockleken och fungerar som motsvarande en proteinsubenhet flera gånger.
Förutom dessa underenheter, som är kända i litteraturen som α -underenheter, har vissa joniska kanaler också en eller flera extra underenheter (ß eller γ) som reglerar öppningen och stängningen av dem.
Specificiteten för varje kanal är relaterad till porens diameter som bildas av de transmembranala proteinerna och sidokedjorna (─r) hos aminosyrorna som gör dem.
På detta sätt finns det kanaler som bara missar natrium, kalium, kalciumjoner och så vidare, eftersom sidokedjorna fungerar som "sikt".
Ytterligare strukturella egenskaper
En annan viktig egenskap hos många kanaler är grindarna. Kanalerna med dessa egenskaper kan öppnas eller stängas framför lokala förändringar som förekommer i membranal mikroenching som omger kanalen.
Beroende på typ av kanal kan dessa förändringar vara mekaniska, termiska (temperaturförändringar), elektriska (spänningsförändringar) eller kemikalier (bindande en ligand).
I de så kallade passiva jonkanalerna, som är de som förblir öppna och tillåter det specifika steget för bestämda joner, har dessa strukturer inte grindar eller är känsliga för ligander eller andra typer av stimuli.
I andra joniska kanaler, som är känsliga för närvaro eller anslutning av ligander, finns det ett bindande ställe för ligand antingen på den extracellulära sidan eller för cellcytosolen och i dessa fall har porerna eller kanalerna en grind som kan öppnas eller stängas Enligt statusen som hans ligand.
Mekanism för sekunder för att öppna eller stängas av kanaler
När det gäller att ha en plats för ligand i den intracellulära delen har dessa kanaler vanligtvis andra budbärare som ligander. Ett exempel på joniska kanaler som öppnar eller stängs av mekanismer för andra budbärare är det för luktreceptorer:
Luktande molekyler binder till sina receptorer på den extracellulära sidan. Dessa receptorer är i sin tur knutna till ett G -protein som aktiveras som i sin tur aktiverar adenylciclas -proteinet som bildar AMPC, vilket är en andra messenger.
AMPC ansluter sig till en intracellulär korsningsplats för kalciumkanaler, vilket resulterar i dess öppnings- och kalciuminträde i cellen.
Som om det var en dominoeffekt, går kalcium till en plats för att länka en annan klorkanal, som genererar dess öppning och utgång av denna jon, vilket orsakar depolarisering av luktcellen.
Det är viktigt att notera att förändringarna som genereras av liganderna eller stimuli som påverkar de joniska kanalerna motsvarar de konformationella förändringarna av proteinerna som utgör kanalens struktur.
Med andra ord, de konformationella förändringarna som kan flytta en grind och stänga eller öppna en kanal är inget annat än tillvägagångssättet eller distansen av proteinsubenheterna som komponerar det.
Andra aktiverings- och inaktiveringsmekanismer
Vissa kanaler, särskilt spänningsberoende kanaler, kan ange ett eldfast tillstånd under vilket samma spänningsförändring som aktiverade dem nu inte längre aktiva dem.
Det kan tjäna dig: Cellbiologi: Historia, vilka studier, applikationer och konceptTill exempel, i spänningsberoende kalciumkanaler, öppnar spänningsförändringen kanalen och kalcium kommer in och, en gång inuti cellen, går samma jon till en gemensam kanalkorsningsplats och den stängs.
En annan form av reversibel inaktivering av kalciumkanalen som förklarar dess eldfasta efter aktivering är parasforylering av kanalen på grund av ökningen av den interna kalciumkoncentrationen.
Det vill säga, en kalciumkanal kan oåterkallelig inaktiveras på grund av närvaron av patologiskt höga koncentrationer av jonen, som förmedlar rekryteringen av utspelande enzymer av andra proteiner aktiverade av kalcium.
De kanaler som regleras av Ligando kan komma in i ett eldfast tillstånd när de förlängs till sin ligand och får denna mekanism namnet på desensibilisering.
Läkemedel, gifter och toxiner kan påverka regleringen av joniska kanaler, stänga dem eller hålla dem öppna eller i vissa fall ockupera ligandplatsen och därmed störa deras funktion.
Funktioner
Joniska kanaler har mångfald av funktioner, direkt eller indirekt.
- De är ansvariga för att reglera flödet av joner genom plasma och organelarmembran i alla celler.
- Tillåt förekomsten av kontroll över de olika jonerna intracellulära koncentrationer.
- I neuroner och i muskelceller kontrollerar joniska kanaler variationerna i membranpotentialen som inträffar under handlingspotentialer och under post -synAptic Post -Synaptic potential för effektorceller.
- Kalciumkanaler som genererar nettokalciumflöden till intracellulärt utrymme är ansvariga för aktiveringen av många enzymer och proteiner som deltar i många metaboliska processer.
- På samma sätt initierar ökningen av kalcium på grund av en ökning av transporten mekanismen för befrielse av neurotransmittorer till det synaptiska utrymmet för neuroner.
- Därför är joniska kanaler också relaterad till cellulära kommunikationsmekanismer.
General för transport genom membranet
Som nämnts ovan kan membranaltransportmekanismerna vara aktiva eller skulder enligt eller inte energin i cellen där de hittas. Passiva mekanismer klassificeras som enkel diffusion och underlättad diffusion.
Enkel diffusion
Enkel diffusion tillåter passage genom fosfolipidstrukturen hos små storlekar fett -lösliga molekyler, med apolära och belastningsfria funktioner.
Således passerar till exempel gaser såsom syre (O2) och koldioxid (CO2), etanol och urea för att nämna några, till förmån för deras koncentrationsgradient.
Underlättad diffusion
Den spridda diffusionen är en som underlättas av protein och denna passiva transportmekanism finns det två typer: jonkanaler och transportproteiner eller transportproteiner.
Joniska kanaler är den mest använda mekanismen av jontransportceller som inte kan passera med enkel diffusion, antingen för att de har elektrisk laddning och membranfosfolipider avvisar dem på grund av deras storlek och polaritet eller någon annan egenskap.
Diffusionen som tillhandahålls av transportproteiner används för transport av större ämnen med eller utan belastning, såsom glukos och andra sockerarter.
Kan tjäna dig: gränssnittActive Membranal Transport är en som inträffar mot koncentrationsgradienten för det lösta ämnet som transporteras och kräver den ATP -formade energiförbrukningen. Bland transportörerna av denna typ är pumpar och vesikulär transport.
Som ett exempel på pumparna är natrium/kalium, som tar tre Sodios och introducerar två kaliumer. Det finns också kalciumpumpar.
https: // giphy.com/gifs/qsib5zeioyRufkuusb
Som ett exempel på vesikulär transport är endocytos, exocytos, pinocytos och fagocytos; Alla dessa aktiva transportmekanismer.
Typer av jonkanaler
Från denna punkt kommer referens att göras till joniska kanaler som tillåter passage av joner genom ett membran till förmån för dess koncentrationsgradienter, det vill säga de är passiva transportkanaler.
Generellt sett är var och en av dessa kanaler specifika för en enda jon, med förbehåll för några få kanaler som tillåter transport av jonpar.
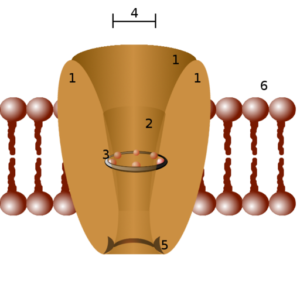 Strukturell schema för en jonisk kanal (källa: utlopp (Paweł Tokarz) vid PL.Wikipedia [Public Domain] via Wikimedia Commons)
Strukturell schema för en jonisk kanal (källa: utlopp (Paweł Tokarz) vid PL.Wikipedia [Public Domain] via Wikimedia Commons) Ett sätt att klassificera joniska kanaler grupperar dem enligt mekanismen som är ansvarig för dess öppning. Således har passiva kanaler, kanaler som regleras av spänning (spänningsberoende), kanaler som regleras av ligando och kanaler som regleras av mekaniska stimuli.
- Passiva kanaler: De är kanaler som är permanent öppna och inte svarar på någon typ av stimulans; Dessa är specifika för vissa joner.
- Spänningsberoende kanaler: Dessa kan öppnas eller stängas (beroende på kanalen) inför förändringar i membranspänningen. De är mycket viktiga för cellsignalering, särskilt i det centrala nervsystemet hos däggdjur.
- Lättberoende kanaler: även kallade kanaler med länkande dörr eller reglerade genom länkning, är allmänt fördelade i de olika kroppscellerna hos människor, men i nervsystemet utgör de de joniska kanaler som aktiveras av neurotransmittorer och är viktiga för synaptisk överföring och intercellulärt skylt.
Exempel på ligandberoende jonkanaler aktiverade av neurotransmittorer är natrium/kaliumkanaler aktiverade av glutamat.
Aktiveringen av kolinergiska receptorer öppnar i detta fall föreningen av acetylkolin till det postsynaptiska membranet (kanalligand), ligandberoende natriumkanaler och tillåter inträde av denna jon efter dess koncentrationsgradient.
- Kanaler reglerade av mekaniska stimuli: De är kanaler som kan aktiveras av distension eller tryck. Dessa mekaniska krafter överförs till kanalen genom cytoskeletten och kanalen öppnas.
Referenser
- Björn, m. F., Connors, b. W., & Paradiso, m. TILL. (Eds.). (2007). Neurovetenskap (Vol. 2). Lippinott Williams & Wilkins.
- Institutionen för biokemi och molekylär biofysik Thomas Jesell, Siegelbaum, S., & Hudspeth, a. J. (2000). Principer för neural vetenskap (Vol. 4, sid. 1227-1246). OCH. R. Kandel, J. H. Schwartz, & t. M. Jesell (Eds.). New York: McGraw-Hill.
- Lewis, c. TILL., & Stevens, c. F. (1983). Acetylkolinkanaljon selektivitet: INSupplev en vattenhaltig miljö. Proceedings of the National Academy of Sciences, 80(19), 6110-6113.
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger principer för biokemi. Macmillan.
- Rawn, j. D. (1998). Biokemi. Burlington, Massachusetts: Neil Patterson Publishers.
- Viana, f., av Peña, och., & Belmonte, c. (2002). Specificitet för kall termotransduktion bestäms av differentiellt jonkanaluttryck. Natur neurovetenskap, 5(3), 254.
- « Flora Cocoid -egenskaper, genrer, patologier, diagnos
- Trachycarpus egenskaper, livsmiljö, distribution, arter »