

Germceller egenskaper, bildning, typer, migration
- 2707
- 479
- PhD. Lennart Johansson
De bakterieceller De är föregångarcellerna i gameterna i dessa djur med sexuell reproduktion. Dessa är embryonala föregångare som skiljer sig från flera somatiska celllinjer (soma = kropp) mycket tidigt i utvecklingen av de flesta arter.
Praktiskt taget uppstår alla organismer som reproducerar sexuellt från sammansmältningen av två gametiska celler. Gameterna är specialiserade celler som bär hälften av den genetiska informationen från individen som producerar dem, fadern och modern (de är haploida celler).
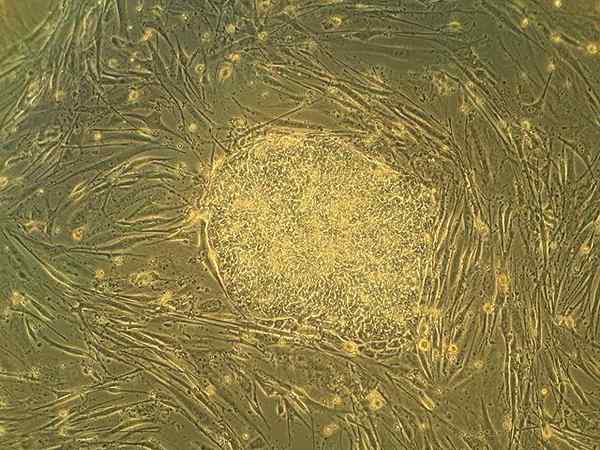 Mänskliga embryonala stamceller i cellkultur. Ryddragyn på engelska Wikipedia [public domain]
Mänskliga embryonala stamceller i cellkultur. Ryddragyn på engelska Wikipedia [public domain] Alla gameter som produceras av ett djur bildas från en speciell cellinje som kallas kimlinjen, som utvecklas enligt en komplex uppsättning specifika signaler. Dessa celler representerar den viktigaste "överförings" -vägen för genomet och de cytosoliska komponenterna från en generation till en annan.
Germceller är ansvariga för specifikations- och evolutionsprocesserna, eftersom det är de förändringar som inträffar i dessa som överförs från en generation till följande. Dessutom är dessa celler det som förmedlar överföringen av föräldrarnas ärftliga sjukdomar till sina barn, särskilt i människan.
[TOC]
Egenskaper hos bakterieceller
Germceller är "pluripotenta" eller "totipotenta" embryonala celler, det vill säga de kan differentieras i nästan alla celltyp under lämpliga förhållanden och signaler. Dessutom är de kompetenta celler för sin "autrenovation", eftersom de är ansvariga för sin egen förnyelse.
Dessa celler är de enda som kan producera gameterna, som är cellerna som kan bilda en ny organisme, egenskaper som de andra cellerna i ett embryo förlorar när de skiljer sig åt.
Vissa författare betraktar dem, som en art "stamceller", eftersom de bildar organ men nya individer. På samma sätt är dessa celler de viktigaste medlen som arter utvecklas och är fordon för överföring av ärftliga sjukdomar, särskilt hos människor.
Germceller producerar gameter genom processer som kallas meios och gametogenes (ovogenes och spermatogenes hos många djur), som är karakteristiska och unika för denna grupp celler.
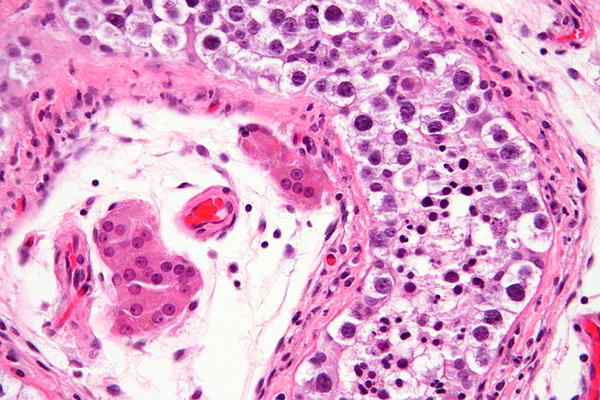
 Seminiferous tubuli med mogen spermier. Nephron [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)]
Seminiferous tubuli med mogen spermier. Nephron [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)] Träning
Germceller skiljer sig tidigt från andra somatiska celllinjer under embryonal utveckling.
Hur händer det i vissa ryggradslösa djur?
 Drosophila melanogaster fruktfluga. Tagen och redigerad från: Sanjay Acharya [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)]
Drosophila melanogaster fruktfluga. Tagen och redigerad från: Sanjay Acharya [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] I många arter, till exempel fruktfluga D. Melanogaster, Dessa celler är bildade av primordiala blostula -celler som har "ärvt" en cytosolisk determinant känd som "bakterieplasm" eller "bakterieplasma", det vill säga mycket speciellt blastomerer.
Denna bakterieplasm innehåller strukturella element och vissa messenger -RNA och under ovogenes och befruktning lider detta.
Blastomererna som har "Germinal Plasma" är uppdelade asymmetriskt, ärver bakterieplasm till en enda dottercell. När embryotet når gastrula -stadiet börjar den ekvivalenta uppdelningen av dessa celler och populationen av primordialceller i kimlinjen expanderar.
Hur händer det hos människan?
Hos däggdjur som människa, å andra sidan, har involveringen av en "kimplasma" inte rapporterats under processen att bilda primordiala bakterieceller, utan snarare bestäms specifikationen av denna linje av cellcellinteraktioner.
Det kan tjäna dig: Cromafin Celler: Egenskaper, histologi, funktionerPrimordiala bakterieceller, under de tidiga stadierna av embryogenes, är belägna i ett slags extra behandlat fack och hos människor inträffar detta nära den tredje utvecklingsveckan.
När linjen med primordialceller har definierats, migrerar de till kvinnliga eller manliga gonader, där ovogenes eller spermatogenesprocesser aktiveras respektive.
Interaktionen mellan primordialceller med somatiska celler av gonader, utöver närvaron av sexuella kromosomer och andra moderfaktorer, är det som definierar bestämningen av kön i den tyska linjen, även om denna process kan variera avsevärt mellan en art och en annan.
Differentiell genuttryck
För att ta den initiala "separationen" av somatiska celler och groddceller är det första som händer ett differentiellt uttryck för gener, eftersom de karakteristiska generna i somatiska linjer i kimlinjen för att starta "programmet" genetiska kimceller.
Under deras bildningsprocess migrerar dessa celler också där de härstammar till den specifika platsen där gonader kommer att bildas, som är den vuxna gamete -producerande vävnaderna.
Cellmigrering uppnås också genom aktivering av en hel migrerande "maskiner" och av olika "guidemekanismer, som har att göra med flera genetiska och epigenetiska faktorer (som har att göra med modifieringen av nukleotidsekvensen).
Migrering
De primordiala bakteriecellerna, de som ger upphov till de "sanna" groddcellerna, bildas långt ifrån där de utvecklas och för att etablera sig måste de migrera till de platser där äggstockarna och testiklarna är belägna, som är kvinnliga och kvinnliga och gonader.
Ursprungligen observeras primordiala bakterieceller under gastrering som ett cellkluster vid basen av allantoiderna, som är ett extraembrionärt membran som bildas som en evagination av det primära matsmältningskanalen i embryot.
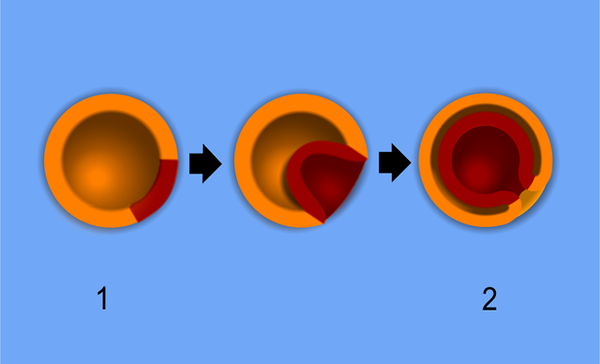 Gastulering av ett diblistiskt djur. 1-blast. 2-gastrula. Pidalka44 [public domain]
Gastulering av ett diblistiskt djur. 1-blast. 2-gastrula. Pidalka44 [public domain] I detta skede förvärvar primordialceller polariserad morfologi och vissa experiment har visat att de sträcker sig långa processer när de mobiliserar.
Därefter blir dessa tydliga i den bakre tarmen och dyker sedan upp från tarmens rygg och migrerar i sidled, koloniserar könsryggarna.
När de ursprungliga cellerna rör sig från den bakre tarmen till den bindväv som omger den, förlängs den senare, och bildar tarmmesenteri (vävnaden som täcker tunntarmen och förenar den med bukväggen), en process som inträffar medan celler dyker upp genom tarmväggen.
Kontrollmekanismer
Ankomsten av prekursorcellerna till gonadalvävnaden styrs av de somatiska cellerna i dessa strukturer, som tydligen utövar en "kemoatracent" -effekt på den första.
Det har visats experimentellt att uttrycket av en gen känd som Fragilis Det har mycket att göra med utvecklingen av rörlighet i primära bakterieceller.
Kan tjäna dig: cytoskelettDenna gen.
Vissa författare anser emellertid att migrationsvägen för dessa celler styrs på miljönivå istället för att vara en autonom process.
I sin passage mot gonaderna och en gång i dem multipliceras dessa celler genom mitos, vilket genererar flera kloner som lyckas öka antalet individer i den cellulära befolkningen.
Bakteriecelltyper
När primära bakterieceller når sina slutliga vävnader utvecklas dessa i kvinnliga bakterieceller eller i manliga bakterieceller, i förekommande fall och de endogena och exogena signalerna som de får.
De kvinnliga gonaderna är äggstockarna och de manliga gonaderna är testiklarna. En gång i dessa vävnader multipliceras primordialceller med hög hastighet, men mönstren för denna mitotiska spridning skiljer sig mellan dem.
Det finns då två typer av bakterieceller kända som ovogonier och spermatogonier.
Kvinnliga bakterieceller: ovogonier
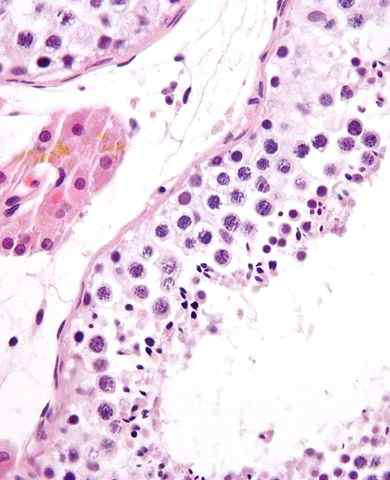
Ovogonier. Källa: Chassot A-A, Gregoire EP, Lavery R, Taketo MM, av Rooij DG, et al. [CC av 3.0 (https: // CreativeCommons.Org/licenser/av/3.0)] Ovogonier är mitotiskt aktiva celler. De är uppdelade intensivt under embryonal utveckling, särskilt från den andra till den femte graviditetsmånaden hos människor, som bildades upp till mer än 7 miljoner av dessa celler, även om vissa är naturligt degenererade.
Dessa celler är inte uppdelade av mitos igen i postnatalstegen, men de skiljer sig regelbundet. Under de sista stadierna av fosterutvecklingen börjar dessa emellertid delas upp av meios, en process som kvarstår i ”arrestering” fram till början av puberteten.
Manliga bakterieceller: Spermatogonier
Proliferationen av spermatogonier skiljer sig något från oogonier, för även om de börjar bilda och multiplicera i de embryonala testiklarna, behåller de sin förmåga att uppdelning i praktiskt taget allt postnatalt liv.
Testiklarnas seminiferösa kanaler avgränsas internt med groddande spermatogonier och några av de populationer som består av dessa celler är uppdelade av mitos. I början av puberteten börjar spermatogoniasgrupper (primära spermatocyter) dela sig genom meios för att träna sekundära spermatocyter som kommer att ge upphov till spermátidas haploid.
Mutationer
Germceller är "fabrikerna" där "fordon" för överföring av informationen från en generation till nästa. På samma sätt är dessa celler av stor betydelse för evolutionära processer, eftersom nästan alla modifieringar som lidits kommer att skrivas ut i avkommorna.
Vi kan säga att DNA från alla celler i en organisme är benägen att mutationer och trots att mutationer i somatiska celler är viktiga i samband med många sjukdomar och andra tillstånd, sträcker sig dessa inte alltid utöver livet i livet för porta individen.
Mutationer i groddlinjen, å andra sidan, bidrar direkt till evolutionära genetiska processer, eftersom dessa förändringar kan gå från en generation till nästa genom gameter och zygote.
Av dessa skäl kan mutationer i bakterieceller endast vara synliga i avkomman och detta beror, mycket ofta, homozygositet eller heterozygositet hos de drabbade generna i varje förälder.
Kan tjäna dig: kromatin: typer, egenskaper, struktur, funktionerOrsakerna till mutationer i groddlinjen är många, eftersom de kan uppstå som svar på endogena eller exogena signaler. Vissa av dessa mutationer producerar sjukdomar som kan ärvas av moderlinjen eller av faderns linje, beroende på fallet.
Tumörer i bakterieceller
Den okontrollerade uppdelningen av cellerna i nästan vilken vävnad som helst i människokroppen, liksom hos andra djur, kan resultera i bildning av tumörer, som kan vara godartade eller maligna.
De som uppstår från bakterieceller kallas vanligtvis neoplasmer och kan vara:
- Grodd
- Teratom
- Embryonkarcinom
- Endodermala sinus tumörer
- Koriokarcinom
Dessa tumörer kan regelbundet förekomma i de inre regionerna i gonaderna, även om de också kan vara relaterade till spridning eller avvikande migration av primordiala bakterieceller, vilket innebär att de kan visas i olika områden i kroppen.
Elektronisk mikrografi av en intratubulär neoplasi av bakterieceller i testiklarna eller "seminom" (Källa: Nephron [CC BY-SA (https: // Creativecommons.Org/licenser/BY-SA/3.0)] via Wikimedia Commons) Tumörer associerade med den primordiala kimcellinjen kallas germinom, medan embryonala karcinom är de som härstammar från embryonala "moder" -celler eller härledda celler.
Usually, primordial germ cells that are formed in extragonadal places are removed, but teratomas are aberrant growths of extragonadal germ cells that have managed to survive, and that are composed of random mixtures of differentiated tissues such as cartilage, skin, hair or teeth.
Endodermala sinus tumörer är de som är bildade från celler härrörande från extrambrionära vävnader och som är differentierade, som bildar endodermal vitellinsäck. Om tumören tvärtom bildas i det trofoblastiska skiktet, kallas detta choriocarcinom.
Tumörerna i groddlinjerna i äggstockarna representerar cirka 20% av alla äggstocks tumörer, är strömmar hos flickor och ungdomar upp till 20 år och nästan alltid om teratom av ond natur.
Bland dessa har utmärkta dysgerminom, som är solida och köttiga tumörer som har en mjuk täckning, sammansatt av aggregat av polygonala utseendeceller, med framträdande plasmamembran och ett stort antal cytosoliska granuler.
Referenser
- Carlson, f. M. (2018). Mänsklig embryologi och utvecklingsbiologi e-bok. Elsevier Health Sciences.
- Jennings, m. T., Gelman, r., & Hochberg, f. (1985). Intracranial bakterie-celllumörer: naturhistoria och patogenes. Journal of Neurosurgery, 63 (2), 155-167.
- Kurman, r. J., & Norris, h. J. (1977). Maligna bakterie tumörer i äggstocken. Human Pathology, 8 (5), 551-564.
- Molyneaux, K., & Wylie, c. (2004). Natm cellmigration naterm. International Journal of Developmental Biology, 48 (5-6), 537-543.
- Hårstrån, e., Foreboscco, a., & Schlessinger, D. (2011). Germcellbildning från eminiska stamceller och användning av somatiska cellkärnor i oocyter. Annals of the New York Academy of Sciences, 1221 (1), 18.
- Richardson, f. OCH., & Lehmann, r. (2010). Mekanismer som riktar NATM -cellmigration: Strategier från olika organisationer. Nature Reviews Molecular Cell Biology, elva(1), 37-49.
- Goren, m. (2010). Cellbiologin i groddcellens livscykel. Nuvarande åsikt i cellbiologi, 22 (6), 707.
- Wylie, c. (1999). Bakterieceller. Cell, 96 (2), 165-174.
- « Extracellulär fluidkomposition och funktioner
- Cellularbiologihistoria, vilka studier, applikationer och koncept »