

Transkriptionsfaktormekanism för handling, typer, funktioner
- 1968
- 425
- Anders Larsson
En Transkriptionsfaktor Det är ett reglerande "tillbehör" -protein som är nödvändigt för genetisk transkription. Transkription är det första steget i genetiskt uttryck och innebär överföring av information som finns i DNA till en RNA -molekyl, som därefter behandlas för att ge upphov till genprodukter.
RNA -polymeras II är enzymet som ansvarar för transkriptionen av de flesta eukaryota gener och producerar, utöver några små RNA, messenger -RNA som sedan kommer att översättas till proteiner. Detta enzym kräver närvaro av en typ av transkriptionsfaktorer som kallas allmänna eller basala transkriptionsfaktorer.
Detta är emellertid inte de enda transkriptfaktorerna som finns i naturen, eftersom det finns "icke-generella" proteiner, både i eukaryoter och i prokaryoter och bågar, som är involverade i regleringen av vävnadsspecifik genetisk transkription (i multicelliga organismer ) eller i regleringen av genaktivitet som svar på olika stimuli.
Dessa transkriptionsfaktorer är effektorer av stor betydelse och kan hittas praktiskt taget i alla levande organismer, eftersom de representerar den viktigaste källan till reglering av genetiskt uttryck.
Detaljerade studier av olika transkriptionsfaktorer i olika typer av levande organismer indikerar att de har en modulär struktur, där en specifik region är ansvarig för interaktion med DNA, medan andra ger stimulerande eller hämmande effekter.
Transkriptionsfaktorerna deltar då i modelleringen av de genetiska uttrycksmönstren som inte har något att göra med förändringarna i DNA -sekvensen, men med epigenetiska förändringar. Vetenskapen som ansvarar för studien av dessa förändringar kallas epigenetik.
[TOC]
Handlingsmekanism
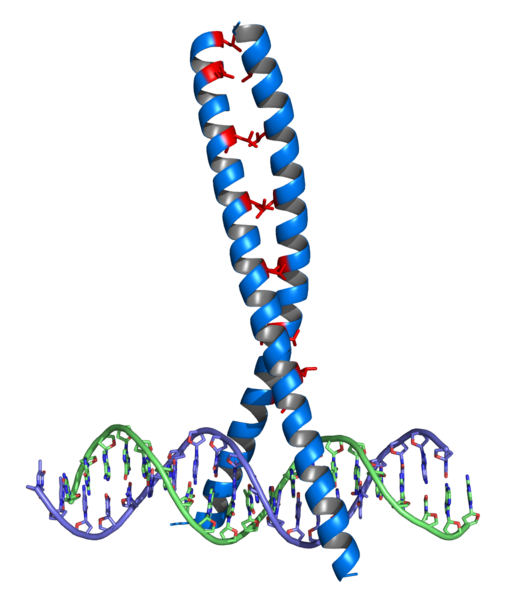
För att utöva sina funktioner måste transkriptionsfaktorer kunna känna igen och gå med specifikt en viss DNA -sekvens för att positivt eller negativt påverka transkriptionen av nämnda region i DNA.
De allmänna transkriptionsfaktorerna, som i princip är desamma för transkription av alla typ II -gener i eukaryoter, monteras först på promotorregionen för genen, vilket riktar placeringen av polymerasenzymet och "öppningen" av dubbelpropellen.
Processen ges av flera på varandra följande steg:
- Förening av den allmänna transkriptionsfaktorn TFIID till en sekvens av upprepad Timina (T) och adenin (A) i genen som kallas ”rutan Tata”; Detta orsakar DNA -distorsion som är nödvändig för föreningen av andra proteiner till promotorregionen.
- Bakre montering av andra allmänna faktorer (tfiib, tfiih, tfih, tfiie, tfiif, etc.) och av RNA -polymeras II, bildar vad som kallas Transkriptionsinitieringskomplex.
Kan tjäna dig: kromatider- Befrielse av initieringskomplexet, fosforylering av polymeras med TFIIH -faktorn och början av transkriptionen och syntesen av en RNA -molekyl från sekvensen för genen som transkriberas.
Aktivering och förtryck av transkription
Som nämnts kan "icke -generella" transkriptionsfaktorer reglera uttrycket av gener, vare sig det är positivt eller negativt.
Aktivering
Vissa av dessa proteiner innehåller, utöver de strukturella domänerna i DNA -unionen, andra skäl som kallas aktiveringsdomäner, som är rika på sura syrarester, glutamin- eller prolinrester.
Dessa aktiveringsdomäner interagerar med elementen i de allmänna transkriptionsfaktorkomplexen eller med relaterade koaktiverande molekyler som direkt interagerar med komplexet. Denna interaktion resulterar i stimulering av sammansättningen av det transkriptiva komplexet eller i ökningen av dess aktivitet.
Undertryckande
De flesta transkriptionsfaktorer hämmar transkription när man stör aktiviteten hos transkriptionsfaktorer som verkar positivt, vilket blockerar dess stimulerande effekt. De kan arbeta genom att blockera föreningen med den positiva faktorn till DNA eller agera på de faktorer som inaktiverar kromatins struktur.
Andra hämmande faktorer agerar direkt som blockerar transkriptionen utan att blockera verkan av någon aktiverande transkriptionsfaktor; och minska basalnivån för transkription, till en ännu lägre nivå än vad som uppnås i frånvaro av aktiverande faktorer.
Liksom att aktivera proteiner fungerar förtryckande faktorer direkt eller indirekt med basala eller allmänna transkriptionsfaktorer.
Grabbar
Även om de flesta transkriptionsfaktorer klassificeras enligt egenskaperna eller identiteten hos deras DNA -fackliga domäner, finns det några, också klassificerade som transkriptionsfaktorer, som inte interagerar direkt med DNA och är kända som transkriptionsfaktorer "indirekt".
Direkta transkriptionsfaktorer
De är de vanligaste transkriptionsfaktorerna. De har DNA -fackliga domäner och kan aktivera eller hämma genuttryck med hjälp av deras förening till specifika DNA -regioner. De skiljer sig från varandra särskilt med avseende på deras DNA -fackliga domäner och deras oligomeriseringstillstånd.
De mest studerade och erkända familjerna av denna typ av faktorer är:
Helix-Gutero-Hélice ("Spiral-helix”, Hth)
Detta var den första familjen av faktorer med DNA -fackliga domäner som upptäcktes och finns i många eukaryoter och prokaryoter. Dess anledning till erkännande består av en α -propell, en tur och en andra propeller a.
De har bevarat domäner av glycin i området i svängen och även ett visst hydrofobt avfall som hjälper till att stabilisera arrangemanget av de två propellerna i HTH -enheten.
Kan tjäna dig: genetisk poolHomeodominium
Det finns i ett stort antal eukaryota reglerande proteiner. De första sekvenserna erkändes i reglerande proteiner för utvecklingen av Drosophila. Denna domän innehåller en HTH-anledning att gå med i DNA och en ytterligare a-propell, utöver en utökad N-terminal arm.
Zinkfingrar
Upptäcktes i transkriptionsfaktorn tfiiia av Xenopus Och det har visats att de deltar i många aspekter av eukaryota genetisk reglering. De finns i proteiner inducerade av signaler om differentiering och tillväxt, i protooncogenes och i vissa allmänna transkriptionsfaktorer.
De kännetecknas av närvaron av repetitioner i ett parti av motiv av zink av 30 avfall som innehåller flera avfall och histidinavfall.
Steroidreceptorer
Denna familj innehåller betydande reglerande proteiner som, förutom att ha en domän för föreningen av hormoner, har en domän av DNA -union och vanligtvis fungerar som transkriptionella aktivatorer.
Unionens domäner innehåller 70 avfall, bland vilka 8 är kvarhållna cysteinrester. Vissa författare anser att dessa faktorer kan bilda ett par zinkfingrar, med tanke på närvaron av två fyra cystein -spel.
Leucina och Helic-Bucle-Herice stängning ("Spiral-loop-helix ")
Dessa transkriptionsfaktorer är involverade i differentiering och utveckling och arbetar för bildandet av en heterodimer. Leucin -stängningsdomänen observeras i olika eukaryoter och kännetecknas av två underdomäner: stängningen av leuciner som mäter dimerisering och en basregion för förening med DNA.
Skälen ß Skäl
De finns främst i eukaryota faktorer och utmärker.
Indirekta transkriptionsfaktorer
Denna typ av transkriptionsfaktorer utövar dess reglerande effekter på genetiskt uttryck inte genom dess direkta interaktion med DNA, utan genom protein-proteininteraktioner med andra transkriptionsfaktorer som interagerar med DNA. Det är därför de kallas "indirekt".
Den första som beskriver var transaktivatorn för viruset "Herpes enkel " (HSV) känd som VP16, som binder till OCT-1-faktorn när cellerna är infekterade med detta virus, vilket stimulerar transkriptionen av en specifik gen.
Faktorerna av denna typ, som de som är bindande till DNA, kan aktivera eller förtrycka gentranskription, så att de kallas "tvång" respektive "korrigerade".
Reglering
Dessa proteiner kan regleras på två nivåer: i sin syntes och i deras aktivitet, vilket beror på olika variabler och flera situationer.
Syntesreglering
Regleringen av dess syntes kan relateras till uttrycksvävnadsspecifik för vissa transkriptionsfaktorer. Ett exempel på detta kan vara MyoD -faktorn, syntetiserad endast i skelettmuskelceller och är nödvändig för att differentieringen av dess odifferentierade fibroblaster.
Kan tjäna dig: länkade generÄven om regleringen av syntes används i grunden för kontroll av genetiskt uttryck i specifika celltyper och vävnader, är detta inte det enda sättet, eftersom syntesen av faktorer involverade i induktion av gener som deltar i svaret också regleras till flera stimuli.
Verksamhetsreglering
En annan regleringsmekanism för transkriptionsfaktorer är regleringen av dess aktivitet, som har att göra med aktiveringen av andra befintliga transkriptionsfaktorer som utövar positiva eller negativa effekter på aktiviteten hos en viss faktor.
Aktiveringen av dessa "sekundära" faktorer sker vanligtvis genom olika mekanismer såsom ligandförening, förändringar i protein-protein, fosforyleringsinteraktioner, bland andra.
Funktioner och betydelse
Transkriptionsfaktorer deltar i olika processer såsom embryonal utveckling, tillväxt och differentiering, cellcykelkontroll, anpassning till fluktuerande miljöförhållanden, underhåll av specifika proteinsyntesmönster för celler och vävnader, etc.
I växter har de till exempel viktiga funktioner i försvar och i svarhändelser på olika typer av stress. Det har fastställts att osteogenes hos djur styrs av transkriptionsfaktorer, liksom många andra differentieringsprocesser för olika cellinjer.
Med tanke på vikten av dessa proteiner i organismer är det inte ovanligt att tro att förändringar i dessa reglerande element kommer att orsaka allvarliga patologiska förändringar.
När det gäller människor kan patologier associerade med transkriptionsfaktorer vara utvecklingsstörningar (på grund av mutationer som producerar inaktivering av transkriptionsfaktorer, till exempel), störningar i hormonella svar eller cancerformer.
Referenser
- Alberts, b., Dennis, b., Hopkin, K., Johnson, A., Lewis, J., Raff, m.,... Walter, s. (2004). Essential Cell Biology. Abingdon: Garland Science, Taylor & Francis Group.
- Iwafuchi-doi, m., & Zaret, K. S. (2014). Pioneer -transkriptionsfaktorer i cellprogrammering. Gener och utveckling, 28, 2679-2692.
- Latchman, D. (1997). Transkriptionsfaktorer: En översikt. Int. J. Biokemi. Cell. Biogus., 29(12), 1305-1312.
- Latchman, D. S. (2007). Transkriptionsfaktorer. Encyclopedia of Life Sciences, 1-5.
- Marie, s. J. (2008). Transkriptionsfaktorer som styr osteoblastogenes. Arkiv för biokemi och biofysik, 473, 98-105.
- Pabo, c., & Sauer, r. T. (1992). Transkriptionsfaktorer: Strukturfamiljer och principer för DNA -erkännande. Annu. Varv., 61, 1053-1095.
- Singh, K. B., Foley, r. C., & Oñate-Sánchez, L. (2002). Transkriptionsfaktorer i växtförsvar och stressrespons. Nuvarande åsikt i växtbiologi, 5, 430-436.
- « Bussegenskaper, mat, reproduktion, livsmiljö
- Klorofyter egenskaper, livsmiljö, reproduktion, mat »