

Myosinegenskaper, struktur, typer och funktion
- 1484
- 199
- Johan Johansson
De Myosin Det är en molekylmotor, av proteinnatur, som kan gå vidare på aktinfilament i cytosolen. Energin som driver förskjutningen av myosin kommer från ATP -hydrolys. På grund av detta definieras myosin vanligtvis som ett mecanokemi -enzym.
I eukaryoter är myosin ett mycket rikligt protein. Det finns olika typer av myosin, som kodas av en familj av gener. I jäst skiljer sig 5 klasser, medan dussintals har beskrivits hos däggdjur.
Källa: David Richfield (användare: Slashme) När du använder den här bilden i externa verk kan det citeras enligt följande: Richfield, David (2014). "Medical Gallery of David Richfield". Wikijournal of Medicine 1 (2). Doi: 10.15347/WJM/2014.009. ISSN 2002-4436. [CC BY-SA 3.0 (http: // Creativecommons.Org/licenser/BY-SA/3.0/]] Myosin har ett brett utbud av funktioner. Myosin I, bredvid aktin, deltar i rörelsen av keratocyter.
Myosin II ger styvhet till plasmamembranet, deltar i cytokinesis och muskelkontraktion. Båda, myosinas I och II, samarbetar med cellmigrering. Myosinas I och V utför transport av vesiklar längs aktinfilament.
[TOC]
Strukturera
I elektroniska mikrografer har den typiska myosinisoformstrukturen tre domäner: huvud, nacke och svans. Genom hydrolys med chimotripsin erhålls ett segment bestående av huvud och nacke, kallad tung meromiosin (HMM) och ett svanssegment, kallad lätt meromiosin (LMM).
Huvudets behärskning är den n-terminala änden av den tunga kedjan, och svansens domän är den lätta kedjans C-terminala.
Myosinklasser kan differentieras med antalet polypeptiska kedjor som komponerar det, och överflödet och lätt kedjeklassen.
Myosin I har en polypeptidkedja, som bildar ett huvud och dess svans saknar alfa -helikidala regioner. Medan myosinas I och V har två polypeptidkedjor och där.
Myosinas I och V har fackliga platser till calmodulin, som reglerar och fixar CA+2, I lätta kedjor. Myosina jag fixade CA+2 I lätta kedjor, men det gör så annorlunda än kalmodulin.
Egenskaper
På mekanisk nivå har myosinas tre egenskaper, nämligen:
- Myosinhuvudet är den motoriska domänen som går framåt med diskreta steg: Föreningen av myosinhuvudet till ett aktintråd, dess lutning och efterföljande separering producerar myosins rörelse. Denna process är cyklisk och beror på ATP.
- Konformationsförändringar: Hydrolysen av en ATP -molekyl är kopplad vid varje steg i en myosinmolekyl, med hjälp av amplifiering och överföringsnivåer. Detta innebär stora konformationella förändringar av myosin.
Kan tjäna dig: radiell symmetriDen första amplifieringsnivån produceras av förlusten av GAMMA-fosfatgruppen i ATP, som möjliggör en omorganisation av strukturelementen på ATP-unionens plats. Denna omorganisation samordnas med strukturella förändringar på platsen för Union to Actin.
Den andra amplifieringsnivån innebär kommunikation av konformationell förändring i den aktiva platsen till strukturella komponenter i terminal karboxyl.
- Riktning: Det har visat sig att min har en polaritet, eller omvänd riktning, mot slutet (+) av aktintråden. Denna slutsats kommer från utjämningsexperimenten i aktinfilamenten med hjälp av fluorescensoptiskt mikroskop.
Funktioner
Myosin, bredvid aktin, deltar i muskelkontraktion, cellulär vidhäftning, cytokinesis, beviljar styvhet till kortikala membran och förskjutningen av vissa vesiklar, bland andra funktioner.
Myosinfel kan producera patologiska tillstånd. Till exempel är defekter i myosinas I och V relaterade, med myopatier från myosin och pigmenteringsstörningar (Griselli -syndrom). Medan störningarna i myosin isoformer vi orsakar förlust av örat.
Muskelsammandragning
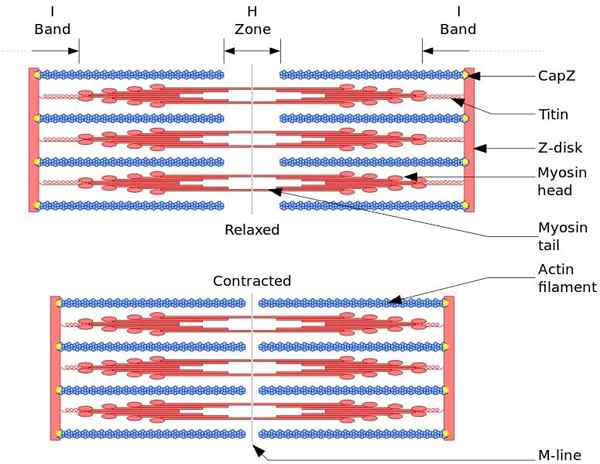
Den funktionella och strukturella enheten i skelettmuskeln är sarkomro. Under muskelkontraktion når sarkomens längd 30% av sin ursprungliga längd.
Sarkomerer består av tjocka, myosin och tunna filament av aktin, som är organiserade på ett komplext sätt. I allmänhet är myosinhuvuden belägna vid filamentets distala ändar och dess svansar mot sarkomero, och organisationen är bipolär.
För att göra muskelkontraktion måste myosinhuvuden, i motsatta ändar, flytta till Z -skivan eller slutet (+) på filamentet. Eftersom organisationen av tjocka filament är bipolär inträffar glidningen av tunna filament på tjocka filament, drivs av ATP.
Förskjutningskraften inträffar eftersom hundratals myosinhuvuden, av ett tjockt glödtråd, interagerar med en tunn filament.
Cytocinesis
Under mitosen, när mikrotubulorna i spindelpolerna är separerade, bildar aktin och myosin II en kontraktil ring i cellen i cellen. Denna ring kontrakterar dess diameter och delar cellen i två delar.
Beviljande av styvhet till kortikala membran
I mutanta celler som saknar myosin II deformeras plasmamembranet lätt när en yttre kraft appliceras. Detta händer eftersom myosin II tillhandahåller aggregeringskraft till plasmamembranproteinerna.
Kan tjäna dig: morfologisk anpassningCell vidhäftning
I epitelvävnaden finns det kontraktila aktin och myosin II -balkar i grannskapen i plasmamembranet och bildar ett cirkulärt bälte som omger cellcellytan. Detta cirkulära bälte bestämmer cellens form och upprätthåller föreningen mellan cellerna.
Kontakten mellan cellerna inträffar av den cirkulära bältesföreningen till celladhesionsmolekyler, med användning av bindande proteiner.
Förskjutning av vissa vesiklar
Experimentella bevis avslöjar att myosin V utför membrantransport från Golgi -apparaten till cellens periferi. Vissa bevis är:
- I nervvävnadsceller, genom immunofluorescens av astrocyter konstaterades att myosin V ligger bredvid Golgi.
- I jäst avbryter mutationer i myosin V -gen proteinsekretion och följaktligen ackumuleras proteiner i cytosol.
- Myosin I -isoformer ansvarar för att transportera vakuoler till cellmembranet. Med hjälp av specifika antikroppar mot myosin I -isoformer konstaterades att dessa isoformer är belägna i olika delar av cellen.
Till exempel, när en levande amoeba är markerad med en antikropp mot myosin IC, arresteras transporten av vakuolen till membranet. På grund av detta expanderar vakuolen och cellen exploderar.
Myosinrelaterade sjukdomar
Myosinas och öronförlust
Det finns många gener och mutationer som producerar öronförlust. Denna sjukdom är ofta monogenetisk .
Mutationer i okonventionella myosinas, med en eller två myosinhuvuden, påverkar det inre örat. Några av de muterade myosin -isforms är myosin IIIA, myosin VIIA och myosin xva. Nyligen upptäcktes två mutationer i myosin VI.
Mutationerna i myosin VI är c.897G> T och P.926Q. Den första mutationen påverkar en region som interagerar med den aktiva belägna, kallad switch i. Homozygot för mutation tidigt uppvisar fenotypen och orsakar allvarliga effekter.
Den andra mutationen påverkar ett belastningsområde med belastning, i en alfa -propell i svansen av myosin VI. Denna region är viktig för den proximala dimeriseringen av motorn och påverkar stereokartfunktionen hos myosin VI.
Det kan tjäna dig: Auxiliary Sciences of BiologyEn annan mutation är P.ASN207SER, som producerar en motor som inte kan producera styrka. Detta beror på att ASN 207 är en aminosyrarest av det aktiva stället, vars funktion är föreningen och hydrolysen av ATP.
Mutationen p.Arg657TRP producerar förlusten av myosin VI -funktionen. Arg -rester är involverad i de konformationella förändringarna som fäster hydrolys till myosinrörelse.
Myosin x och cancer
Myosina x (myo10) är ett okonventionellt myosin som uttrycks i hjärnan, endotelios och många epitelier. Myo10 och tre typer av aktinbaserade projektioner (filopoder, fakturider och prognoser som liknar filopodianer) arbete under cancermetastaser.
Invasiva cancerceller har ett stort antal filopoder och uttrycker höga fascinnivåer. Detta protein utför korsning mellan aktinfilament. För att fly från den primära tumören bildas invaddies, rika på proteolytisk aktivitet, som smälter den omgivande extracellulära matrisen.
När cellerna når den extracellulära matrisen hjälper prognoserna som liknar Philopodes att sprida och kolonisera. Höga nivåer av myo10 indikerar hög aggressivitet och metastas i bröstcancer.
Myoxs tystnad ger förlust av den metastatiska karaktären hos celler, som inte kan bilda aktinbaserade projektioner. Alla dessa prognoser har förståelse -baserade vidhäftningar, som transporteras av MY1010 inom filopodiet.
Myox är involverad i bildandet av centra. Frånvaron av myox gynnar bildningen av multipolära spikar. Myox är också involverad i signalering i cancerceller. Till exempel aktiveras Myox av 3,4,5, -inositol trifosfat (PIP3).
Referenser
- Alberts, b., Johnson, A., Lewis, J., och kål. 2007. Biologi av cellmolekylen. Garland Science, New York.
- Brownstein, Z., Abu-Rayyan, a., Karfunkel-don, D., Sirigu, s., Davido, f., Shohat, m., Frydman, m., Houdusse, a., Kanaan, m., Avraham, K. 2014. Nya myosin -mutationer för ärftlig hörselnedsättning som är uppsvingad genom riktad genomisk fångst och massivt parallell sekvensering. European Journal of Human Genetics, 22: 768 -775.
- Courson, D.S. och Cheney, r.OCH. 2015. Myosin-x och sjukdom. Experimentell cellforskning, 334: 10-15.
- Lodish, h., Berk, A., Zipurski, s.L., Matsudaria, s., Baltimore, D., Darnell, J. 2003. Cellulär och molekylärbiologi. Redaktör Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Mexiko, Sāo Paulo.
- Schliwa, m. och woehlke, g. 2003. Molekylmotorer. Nature, 422: 759 - 765.
- Till värd.D. 2003. Molekylär verktygslåda för intacellulär transport. Cell, 112: 467-480.
- « Exonukleasegenskaper, struktur och funktioner
- Bromo -historia, struktur, elektronisk konfiguration, egenskaper, användningar »