

Mitosfaser, egenskaper, funktioner och organismer
- 4990
- 138
- Hans Olsson
De mitos Det är en celldelningsprocess, där en cell producerar genetiskt identiska dotterceller; För varje cell genereras "döttrar" med samma kromosomal belastning. Denna uppdelning utförs i de somatiska cellerna i eukaryota organismer.
Denna process är ett av stadierna i cellcykeln för eukaryota organismer, som förstås i fyra faser: S (DNA -syntes), M (celldelning), G1 och G2 (mellanliggande faser där mRNA och proteiner inträffar). Tillsammans betraktas faser G1, G2 och S som ett gränssnitt. Den nukleära och cytoplasmiska uppdelningen (mitos och cytokinesis) utgör det sista steget i cellcykeln.
Mydosöversikt. Källa: Viewaprabha [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)] På molekylnivå initieras myitos genom aktivering av ett kinas (protein) som kallas MPF (mognad främjande faktor) och den därmed fosforylering av ett betydande antal cellkomponenter komponenter. Det senare gör det möjligt för cellen att presentera de morfologiska förändringar som krävs för att genomföra uppdelningsprocessen.
Mitos är en asexuell process, eftersom förfädercellen och dess döttrar har exakt samma genetiska information. Dessa celler är kända som diploida genom att bära den kompletta kromosomala belastningen (2N).
Meios, å andra sidan, är processen för celldelning som resulterar i sexuell reproduktion. I denna process replikerar en diploid stamcell dess kromosomer och delar sedan två gånger i rad (utan att replikera dess genetiska information). Slutligen genereras 4 dotterceller med endast hälften av den kromosomala belastningen, som kallas haploider (N).
[TOC]
Generaliteter av mitos
Mitos i encellulära organismer producerar i allmänhet dotterceller som är mycket lik sina föräldrar. Däremot, under utvecklingen av multicelliga varelser, kan denna process orsaka två celler med vissa olika egenskaper (trots att de är genetiskt identiska).
Denna celldifferentiering ger upphov till de olika celltyperna som utgör de multicellulära organismerna.
Under en organisms liv inträffar cellcykeln kontinuerligt och bildar ständigt nya celler som i sin tur växer och förbereder sig för att dela genom myitos.
Cellulär tillväxt och uppdelning regleras av mekanismer, såsom apoptos (programmerad celldöd), som gör det möjligt att upprätthålla en balans och undvika överskott av vävnader. På detta sätt säkerställs det att defekta celler ersätts av nya celler, enligt organisismens krav och behov.
Vilken relevans har denna process?
Förmågan att reproducera är en av de viktigaste egenskaperna hos alla organismer (från encellulär till multicellulär) och cellerna som komponerar den. Denna kvalitet gör det möjligt att säkerställa kontinuiteten i din genetiska information.
Förståelsen av processerna för mitos och meios har haft en grundläggande roll för att förstå de spännande cellegenskaperna hos organismer. Till exempel egenskapen att hålla antalet kromosomer från en cell till en annan inom en individ och mellan individer av samma art.
När vi drabbas av någon typ av snitt eller sår i vår hud, observerar vi hur den skadade huden på några dagar återvinns. Detta inträffar tack vare myitosprocessen.
Faser och dess egenskaper
I allmänhet följer mytos samma processsekvens (faser) i alla eukaryota celler. I dessa faser förekommer många morfologiska förändringar i cellen. Bland dem kondensationen av kromosomerna, bristen på kärnmembranet, separationen av cellen från den extracellulära matrisen och andra celler och uppdelningen av cytoplasma.
I vissa fall betraktas kärnkraftsdelningen och cytoplasmisk uppdelning som olika faser (mitos respektive cytokinesis).
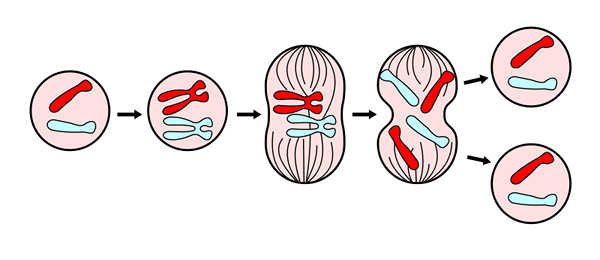
För en bättre studie och förståelse av processen har sex (6) faser utsetts, kallas: propase, utlovat, metafas, anafas och telofas, sedan betraktar cytokinesis som en sjätte fas, som börjar utvecklas under anafas.
 Telofas är den sista fasen av mitos. Hämtad från https: // commons.Wikimedia.Org/wiki/fil: mitosepanel.Jpg. Via Wikimedia Commons
Telofas är den sista fasen av mitos. Hämtad från https: // commons.Wikimedia.Org/wiki/fil: mitosepanel.Jpg. Via Wikimedia Commons Dessa faser har studerats sedan 1800 -talet genom ljusmikroskopet, så idag är de lätt att känna igen enligt de morfologiska egenskaperna hos cellen, såsom kromosomal kondensation, och bildandet av den mitotiska spindeln.
Profas
 Profas. Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons
Profas. Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons Profas är den första synliga manifestationen av celldelning. I den här fasen kan du se utseendet på kromosomer som urskiljbara former på grund av den progressiva komprimeringen av kromatin. Denna kondensation av kromosomer börjar med fosforylering av histona H1 -molekyler av MPF -kinas.
Kondensationsprocessen består av sammandragning och därför minskningen av kromosomernas storlek. Detta inträffar på grund av rullning av kromatinfibrer, vilket ger lättare skördbara strukturer (mitotiska kromosomer).
De kromosomer som tidigare duplicerats under perioden av cellcykeln, förvärvar ett dubbelfilamentutseende, kallad systerkromatider, dessa filament förblir förenade genom en region som heter Centromero. I denna fas försvinner också nukleoli.
Det kan tjäna dig: Cromafin Celler: Egenskaper, histologi, funktionerMitotisk spindelbildning
Av Silvia3 [GFDL (http: // www.gnu.Org/copyleft/fdl.html) eller cc by-sa 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons Under profaset bildas den mitotiska spindeln, bestående av mikrotubuli och proteiner som utgör en uppsättning fibrer.
När spindeln bildas, är mikrotubulierna i cytoskeletten (genom deaktivering av proteinerna som upprätthåller deras struktur), vilket ger det nödvändiga materialet för bildandet av nämnda mitotiska spindel.
Centerna (en organell utan membran, funktionell i cellcykeln), duplicerade i gränssnittet, fungerar som monteringsenheten för spindelens mikrotubulor. I djurceller har centreringen i mitten, ett par centrioler; Men dessa är frånvarande i de flesta växtceller.
Duplicerade centra, de börjar separera det ena från den andra medan mikrotubulierna i spindeln monteras i var och en av dem, börjar migrera till motsatta ändar av cellen.
I slutet av profasen börjar bristen på kärnkraftsförpackningen, förekommer i separata processer: MAS -brottet av kärnkraftsporen, kärnkraftsbladet och kärnmembranen. Denna paus gör det möjligt för mitotisk spindel och kromosomer att börja interagera.
Löfte
Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] I detta skede har kärnhöljet varit helt fragmenterat, så mikrotubulierna i spindeln invaderar detta område och interagerar med kromosomer. De två centra har separerat, var och en i polerna i den mitotiska spindeln, i motsatta ytterligheter av cellerna.
Nu inkluderar den mitotiska spindeln mikrotubulierna (som sträcker sig från varje centrering till mitten av cellen), centren och ett par Osteres (strukturer med radiell fördelning av korta mikrotubulor, som distribueras från varje centrering).
Kromatiderna utvecklade var och en, en specialiserad proteinstruktur, kallad Cinetocoro, belägen i centromeren. Dessa knetocoros är belägna i motsatta riktningar och vissa mikrotubulor följs, kallade mikrotubuli av cinnetocoro.
Dessa mikrotubulor som är fästa vid cinetocoro börjar flytta kromosomen från slutet som de sträcker sig; några från en pol och andra av motsatt pol. Detta skapar en "drag- och krympningseffekt som vid stabilisering gör det möjligt för kromosomen att avsluta mellan cellens ändar.
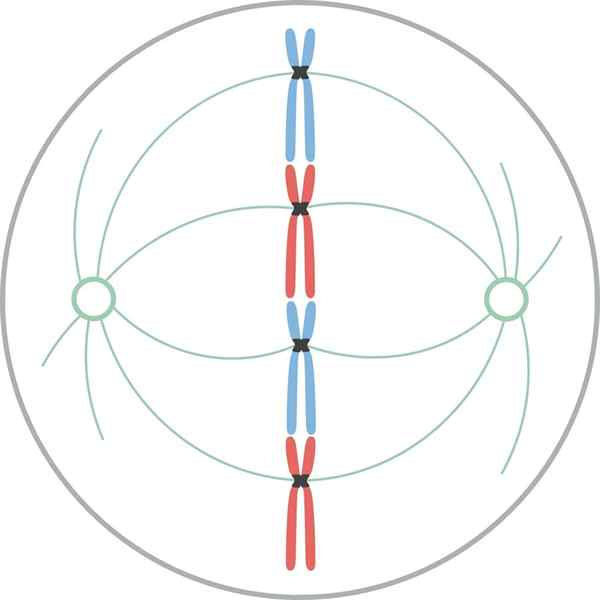
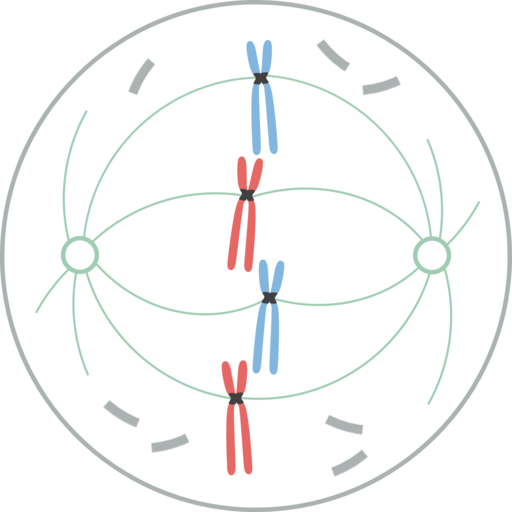
Metafas
Kromosomer inriktade i cellens ekvatorialplatta under mytiskt metafas I metafas är centren belägna i motsatta ändar av cellerna. Spindeln visar en tydlig struktur, i vars centrum kromosomerna finns. Centromererna av dessa kromosomer är fixerade till fibrerna och justeras i ett imaginärt plan som kallas metafasisk plack.
Chromatid cipnetocoros förblir fästa vid mikrotubulierna i Cinetocoro. Mikrotubulor som inte följer knetocoros och sträcker sig från motsatta poler i spindeln, interagerar nu med varandra. Vid denna tidpunkt är mikrotubulierna från osteres i kontakt med plasmamembranet.
Denna tillväxt och interaktion mellan mikrotubuli, slutför strukturen för den mitotiska spindeln och ger ett utseende "Bird Cage".
Morfologiskt är denna fas den som verkar mindre förändringar, så den betraktades som en vilopas. Även om de inte är lätt märkbara förekommer många viktiga processer i den, förutom att de är det längsta stadiet av mitos.
Anafas
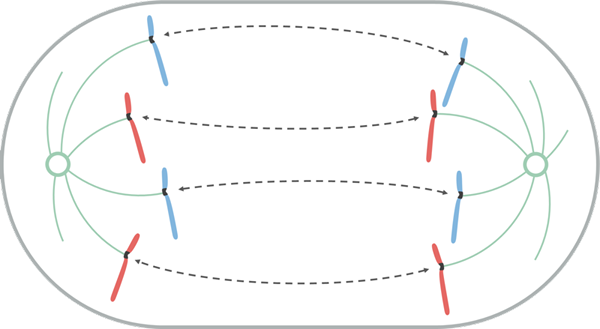 Källa: Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons
Källa: Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons Under anafaset börjar varje par kromatider separera (genom inaktivering av proteinerna som håller dem ihop). Separata kromosomer rör sig mot motsatta ändar av cellen.
Denna migrationsrörelse beror på att mikrotubulierna i Catocoro de Acortan, vilket genererar en "drag" -effekt som gör att varje kromosom rör sig, från sin centromero. Beroende på platsen för centromeren i kromosomen kan det ta under dess förskjutning en viss form som V eller J.
Mikrotubulorna vidhäftade inte till Knetocoro, växer och förlängs genom tubulin vidhäftning (protein) och genom verkan av motorproteiner som rör sig på dem, vilket gör att kontakten mellan dem kan stoppa. När de rör sig bort från varandra gör spindelstolpar också och förlänger cellen.
I slutet av denna fas är kromosomgrupperna belägna i motsatta ändar av den mitotiska spindeln, så varje ände av cellen är med en fullständig och motsvarande uppsättning kromosomer.
Telofas
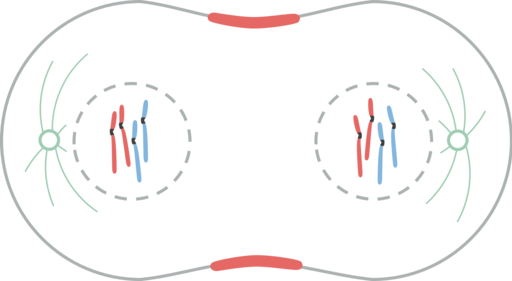 Telofas. Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)]
Telofas. Leomonaci98 [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] Telofas är den sista fasen av kärnkraftsdelningen. Cinetocoro -mikrotubuli sönderdelas medan polära mikrotubuli är ytterligare förlängda.
Det kärnkraftsmembranet börjar bildas runt varje kromosom -spel med hjälp av kärnomslaget i modercellen, som var som cytoplasma vesiklar.
I detta skede är kromosomerna som finns i cellpolerna helt avskräckta på grund av defosforylering av histonmolekyler (H1). Bildningen av elementen i kärnmembranet riktas av flera mekanismer.
Under anafaset började många av de fosforylerade proteinerna vara defosforilerade i profaset. Detta tillåter början av telofasen, kärnvesiklarna börjar återmontera, associerade med kromosomernas yta.
Kan tjäna dig: kondrocyter: egenskaper, histologi, funktioner, odlingÅ andra sidan är kärnkraftsporeen återmonterad som tillåter pumpning av kärnproteiner. Kärnlamina proteiner är defosforylerade, vilket gör att de kan associeras igen för att slutföra bildandet av nämnda kärnlamina.
Slutligen, efter att kromosomerna är helt avskräckta, startas syntesen av RNA, bildar kärnan igen och därmed slutför bildandet av de nya gränssnittskärnorna hos dotterceller.
Cytokinesis

Cytocinesis tas som en händelse åtskild från kärnkraftsdelningen, och vanligtvis i typiska celler åtföljer cytoplasmatisk uppdelningsprocess varje myitos, som börjar i anafas. Flera studier har visat att i vissa embryon inträffar flera kärnkraftsavdelningar före den cytoplasmiska uppdelningen.
Processen börjar med utseendet på ett spår eller klyftan som är markerad i planet för den metafasiska placken, vilket säkerställer att uppdelningen inträffar mellan kromosomgrupperna. Platsen för klyftan indikeras av den mitotiska spindeln specifikt, mikrotubulorna i Ásteres.
I den markerade klyftan finns en serie mikrofilament som bildar en ring riktad till den cytoplasmiska sidan av cellmembranet, till stor del sammansatt av aktin och myosin. Dessa proteiner interagerar med varandra vilket möjliggör sammandragning av ringen runt spåret.
Denna sammandragning genereras av glidningen av filamenten i dessa proteiner, när de interagerar med varandra, på samma sätt som de gör till exempel i muskelvävnader.
Ringens sammandragning fördjupas genom att utöva en "klämningseffekt som slutligen delar upp modercellen, vilket möjliggör separering av dotterceller, med dess cytoplasmiska innehåll i utvecklingen.
Cytocinesis i växtceller
Växtceller har en cellvägg, så deras cytoplasmiska uppdelningsprocess skiljer sig från den som beskrivs tidigare och börjar i telofas.
Bildningen av en ny cellvägg börjar montera mikrotubulierna i den återstående spindeln, som utgör Framoplasten. Denna cylindriska struktur bildas av två mikrotubulsspel som ansluter i dess ändar, och vars positiva poler är inbäddade i en elektronisk plack i ekvatorialplanet.
Små vesiklar från Golgi -apparaten, full av föregångare till cellväggen, rör dig genom mikrotubulierna i Framoplasten till ekvatorialregionen, som kombineras för att bilda en cellplack. Vesiklarnas innehåll är segregerat på denna platta när den växer.
Denna platta växer och slås samman med plasmamembranet längs cellomkretsen. Detta inträffar på grund av den ständiga omorganisationen av mikrotubulierna i Framoplasten i plattans periferi, vilket gör att fler vesiklar kan röra sig mot detta plan och tömma dess innehåll.
På detta sätt inträffar den cytoplasmiska separationen av dotterceller. Slutligen gör innehållet på cellplattan bredvid cellulosa -mikrofibrerna inuti den att slutföra bildandet av den nya cellväggen.
Funktioner
Mitos är en divisionsmekanism i celler och är en del av en av faserna i cellcykeln i eukaryoter. På ett enkelt sätt kan vi säga att huvudfunktionen i denna process är reproduktionen av en cell i två dotterceller.
För encelliga organismer betyder celldelning att generera nya individer, medan för multicelliga organismer är denna process en del av tillväxten och korrekt funktion av hela kroppen (celldelningen genererar utveckling av vävnader och underhåll av strukturer).
Mytosprocessen aktiveras enligt organismens krav. Hos däggdjur, till exempel, börjar röda blodkroppar (erytrocyter) dela fler celler när kroppen behöver en bättre syrefångst. På liknande sätt reproducerar vita blodkroppar (leukocyter) när det är nödvändigt att bekämpa en infektion.
Däremot saknar vissa specialiserade djurceller praktiskt taget mitosprocessen eller är mycket långsam. Exempel på detta är nervceller och muskelceller).
I allmänhet är de celler som är en del av organismens bindande och strukturella vävnad och vars reproduktion endast är nödvändig när en cell har viss defekt eller försämring och måste bytas ut.
Tillväxtreglering och celldelning.
Celltillväxt- och celltivisionssystemet är mycket mer komplicerat i multicelliga organismer än i de encelliga. I det senare är reproduktionen i princip begränsad av tillgången på resurser.
I djurceller arresteras uppdelningen tills det finns en positiv signal som aktiverar denna process. Denna aktivering kommer i form av kemiska signaler från angränsande celler. Detta gör det möjligt att förhindra obegränsad tillväxt av vävnader och reproduktion av defekta celler, vilket allvarligt kan skada organismens liv.
Kan tjäna dig: basmembran: egenskaper, struktur och funktionerEn av mekanismerna som kontrollerar cellmultiplikation är apoptos, där en cell dör (på grund av produktion av vissa proteiner som aktiverar självförstörelse) om den har betydande skador eller är infekterade av ett virus.
Det finns också reglering av cellutveckling genom hämning av tillväxtfaktorer (såsom protein). Således förblir cellerna i gränssnittet utan att fortsätta till m -fasen i cellcykeln.
Organismer som utför det
Mitosprocessen utförs i den stora majoriteten av eukaryota celler, från encelliga organismer som jäst, som använder den som en asexuell reproduktionsprocess, till komplexa multicellulära organismer som växter och djur och djur.
Även om cellcykeln i allmänhet är densamma för alla eukaryota celler, finns det anmärkningsvärda skillnader mellan encelliga och multicelliga organismer. I det förstnämnda gynnas tillväxten och uppdelningen av celler av naturligt urval. I flercelliga organismer begränsas spridningen av strikta kontrollmekanismer.
I encelliga organismer inträffar reproduktion på accelererad, eftersom cellcykeln fungerar ständigt och dotterceller går snabbt mot myitos för att fortsätta med den cykeln. Medan cellerna i multicellulära organismer tar betydligt mer tid att växa och dela.
Det finns också vissa skillnader mellan de mitotiska processerna för växt- och djurceller, som i några av faserna i denna process, men i princip fungerar mekanismen på liknande sätt i dessa organismer.
Celldivision i prokaryota celler
Prokaryot cell I allmänhet växer prokaryota celler och är uppdelade med en snabbare hastighet än eukaryota celler.
Organismer med prokaryota celler (vanligtvis encellulär eller i vissa multicellulära fall) saknar ett kärnmembran som isolerar det genetiska materialet i en kärna, så det är spridd i cellen, i ett område som kallas nukleoid. Dessa celler har en huvudcirkulär kromosom.
Celldelning i dessa organismer är då mycket mer direkt än i eukaryota celler, saknar den beskrivna mekanismen (mitos). I dem utförs reproduktion av en process som kallas binär fission, där DNA -replikation börjar på ett specifikt cirkulärt kromosomställe (ursprung för replikering eller oric).
Två ursprung bildas som migrerar till motsatta sidor av cellen när replikation inträffar, och cellen sträcker sig tills den når dubbelt så stor som storlek. I slutet av replikationen växer cellmembranet in i cytoplasma och delar upp förfädercellen i två döttrar med samma genetiska material.
Mitosutveckling
Utvecklingen av eukaryota celler förde med sig ökningen av komplexiteten i genomet. Detta antydde utvecklingen av mer detaljerade divisionsmekanismer.
Vad som föregick mitosen?
Det finns hypoteser som föreslår att bakteriell uppdelning är föregångsmekanismen för mitos. Ett visst samband mellan proteinerna associerade med binär fission har hittats (vilket kan vara de som förankrar kromosomerna till specifika platser i döttrarnas plasmamembran) med tubulin och aktin i eukaryota celler.
Vissa studier indikerar vissa särdrag i uppdelningen av moderna unicellulära protister. I dem förblir kärnmembranet intakt under mitos. Replikerade kromosomer förblir förankrade i vissa platser i detta membran, som skiljer sig när kärnan börjar sträcka sig under celldelning.
Detta visar en viss slump med den binära fissionsprocessen, där replikerade kromosomer är fixerade på vissa platser i cellmembranet. Hypotesen föreslår sedan att protisterna som presenterar denna kvalitet under sin celldelning, kunde ha upprätthållit detta kännetecken för en förfädercellcell.
För närvarande, förklaringar av varför eukaryota celler av multicellulära organismer ännu inte har utvecklats, är det nödvändigt att kärnmembranet sönderdelas under celldelningsprocessen.
Referenser
- Albarracín, a., & Telulón, till. TILL. (1993). Cellteori under 1800 -talet. Akalutgåvor.
- Alberts, b., Johnson, A., Lewis, J., Raff, m., Roberth, K., & Walter, s. (2008). Biologi av cellmolekylen. Garland Science, Taylor och Francis Group.
- Campbell, n., & Reece, J. (2005). Biologi 7th Upplaga, AP.
- Griffiths, a. J., Lewontin, r. C., Miller, J. H., & Suzuki, D. T. (1992). Introduktion till genetisk analys. McGraw-Hill Inter-American.
- Karp, G. (2009). Cell- och molekylärbiologi: begrepp och experiment. John Wiley & Sons.
- Lodish, h., Darnell, J. OCH., Berk, A., Kaiser, c. TILL., Krieger, m., Scott, m. P., & Matsudaira, s. (2008). Mollecular cellbiologi. Macmillan.
- Segura-Valdez, M. D. L., Cruz-gómez, s. D. J., López-cruz, r., Zavala, g., & Jiménez-García, L. F. (2008). Visualisering av mitos med atomkraftmikroskopet. Dricks. Specialiserad tidning i kemikali-biologiska vetenskaper, 11 (2), 87-90.