

Proteiner g struktur, typer och funktioner
- 4143
- 531
- Karl Johansson
De G antingen Guanin nukleotidbindande proteiner, De är proteiner associerade med plasmamembranet som tillhör en familj av "kopplingar" av signaler som har viktiga funktioner i många signaltransduktionsprocesser i eukaryota organismer.
I litteratur beskrivs G -proteiner som binära molekylära switchar, eftersom deras biologiska aktivitet bestäms av förändringarna i deras struktur som ges av nukleotidarten till vilka de kan förenas: guanosin nukleotider (difosfat (BNP) och typosfär (GTP))).
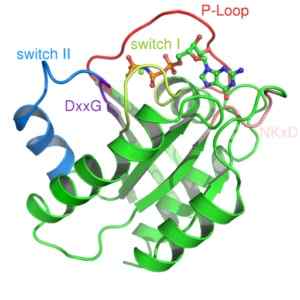
Struktur av RAS -proteinet, ett monomerat G -protein (källa: Mark 'Absturz' [public domain] via Wikimedia Commons) De aktiveras vanligtvis av receptorer från en familj av protein som kallas GPCR -kopplade receptorer (GPCR, engelska "G-protein-kopplade mottagare "), som får en extern initial signal och gör den konformationella förändringar som utlöser aktivering, vilket därefter översätter till aktiveringen av ett annat effektorprotein.
Vissa författare anser att de kodande generna för denna familj av protein utvecklades i duplicering och divergens av en förfäder gemensam gen, vars produkt förfinades och specialiserat mer och mer.
Bland de stora variationerna av cellulära funktioner som dessa proteiner har är translokation av makromolekyler under proteinsyntes, transduktion av hormonella signaler och sensoriska stimuli, liksom regleringen av proliferation och celldifferentiering.
Två klasser har beskrivits från denna typ av protein: små G -proteiner och heterotrofimeriska G -proteiner. De första tre dimensionella strukturen för ett G -protein erhölls mer än ett decennium från ett litet G -protein känt som RAS.
[TOC]
Strukturera
Strukturellt sett erkänns två typer av G -proteiner: små G -proteiner och heterotrimiska G -proteiner, mycket mer komplexa.
Små G-proteiner bildas av en enda polypeptid av cirka 200 aminoaceöst avfall och cirka 20-40 kDa, och i dess struktur finns en bevarad katalytisk domän (domänen G) som består av fem a-propeller, sex p-veckade ark och fem polypeptid slingor.
Heterotrofimeriska G -proteiner är å andra sidan omfattande membranproteiner som består av tre polypeptidkedjor, kända som a, p och y -subenheterna.
-A -underenheten väger mellan 40 och 52 kDa, har en guanin nukleotidbindande region och har GTPASA -aktivitet för att hydrolysera kopplingar mellan GTP -fosfatgrupper.
A -underenheterna för olika G -proteiner delar vissa strukturella domäner såsom GTP -union och hydrolys, men är mycket olika i receptorbindningsställena och effektorproteiner.
Kan tjäna dig: mitotisk spindel-P -underenheten har en något lägre molekylvikt (mellan 35 och 36 kDa).
-Y -underenheten är å andra sidan mycket mindre och har en ungefärlig molekylvikt på 8 kDa.
All heterotrofimerisk g. Dessa två domäner är så starkt associerade att de övervägs som en enda funktionell enhet.
Grabbar
Som nämnts tidigare finns det två typer av G -proteiner: små och heterotriméricas.
Små G -proteiner har funktioner i celltillväxt, proteinsekretion och interaktion mellan intracellulära vesiklar. För sin del är heterotrofimeriska G -proteiner associerade med signaltransduktion från ytreceptorer och fungerar också som switchar som växlar mellan två tillstånd beroende på tillhörande nukleotid.
Små G -proteiner
Dessa proteiner kallas också små GTPAS, små GTP -korsningsproteiner eller Ras -proteinsuperfamilj och bildar en oberoende superfamilj inom den stora klassen hydrolas GTP med reglerande funktioner.
Dessa proteiner är mycket olika och kontrollerar flera cellprocesser. De kännetecknas av en bevarad bindningsdomän till GTP, domänen "G". Föreningen mellan detta nukleotidfosfat orsakar små konformationella förändringar i dess katalytiska domän i GD -proteiner.
Dess aktivitet är intimt relaterad till GTPASAS -aktiverande proteiner (Gap, engelska "GTPase aktiverande protein ") och med Guanina nukleotidutbytesfaktorer (GEF, engelska "Guanin nukleotidutbytesfaktor ”)).
I eukaryoter har fem klasser eller familjer med små G -proteiner beskrivits:
-Raseri
-Rod
-Rab
-SAR1/ARF
-Sprang
Ras och Rho -proteiner kontrollerar genetiskt uttryck och Rho -proteiner modulerar också omorganisationen av cytoskeletten. Rab och SAR1/ARF -gruppproteiner har påverkan på vesikulär transport och proteiner reglerar kärntransport och cellcykel.
Heterotrofimeriska G -proteiner
Denna typ av protein förtjänar också en associering med två andra proteinfaktorer, så att signalvägen från utsidan till den cellulära inre består av tre element i följande ordning:
- De kopplade receptorer En proteiner g
- De G
- De Proteiner eller den Effektiva kanaler
Det finns en stor mångfald av heterotrofimeriskt G -protein och detta är relaterat till den stora mångfalden av α -underenheter som finns i naturen, där endast 20% av aminosyrasekvensen bevaras.
Vanligtvis identifieras heterotrofimeriska G -proteiner tack vare mångfalden av a -underenhet, baserat främst på deras funktionella och sekvenslikheter.
A -underenheterna utgör fyra familjer (GS -familjen, GI/O -familjen, GQ -familjen och G12 -familjen). Varje familj består av en annan "isotyp" som tillsammans lägger till mer än 15 olika sätt av a -underenheter.
Familj gs
Denna familj innehåller representanter som också deltar i den positiva regleringen av cyklasadenylatproteiner och uttrycks i de flesta celltyper. Det består av två medlemmar: GS och Golf.
Abonnemanget "S " Det hänvisar till stimulering (från engelska "Stimulans ") och abonnemanget "Olf " Det hänvisar till "lukt" (från engelska "Olfaction"). Golfproteiner uttrycks särskilt i de sensoriska neuronerna som är ansvariga för lukten.
Familj gI/O
Detta är den största och mångfaldiga familjen. De uttrycks i många celltyper och förmedlar den mottagningsberoende hämningen av flera typer av adenylciclasses (abonnemanget "Yo " avser hämning).
Proteiner med grupp a -underenheter uttrycks särskilt i celler i centrala nervsystemet och har två varianter: A och B.
Familj gq
Proteiner med denna familj av α -underenhet är ansvariga för regleringen av fosfolipas C. Denna familj består av fyra medlemmar vars α -underenheter uttrycks av olika gener. De finns rikligt i leverceller, i njurceller och lungor.
Familj g12
Denna familj uttrycks allestädes närvarande i organismer och det är inte känt med säkerhet vad exakt de cellulära processer som regleras genom proteiner med dessa underenheter är.
P- och y -underenheter
Även om mångfalden av alfastrukturer är avgörande för identifiering av heterotimeriska proteiner, finns det också mycket mångfald med avseende på de andra två underenheterna: Beta och Gamma.
Funktioner
G -proteiner deltar i "kanaliseringen" av signalerna från receptorerna i plasmamembranet till effektorkanalerna eller enzymerna.
Det vanligaste exemplet på funktionen av denna typ av protein är i regleringen av enzymet adenylatcyklasa, enzym som är ansvarigt för syntesen av adenosin 3 ', 5'-monofosfat eller helt enkelt cyklisk AMP, en molekyl som har viktiga funktioner som andra messenger I många kända cellprocesser:
Kan tjäna dig: caliciform celler-Selektiv proteinfosforylering med specifika funktioner
-Genetisk transkription
-Omorganisation av cytoskelettet
-Utsöndring
-Membran depolarisation
De deltar också indirekt i regleringen av inositolerna som signalerar vattenfallet (fosfatidylinitol och deras fosforylerade derivat), som är ansvariga för kontroll av kalciumberoende processer såsom kemiotax och utsöndring av lösliga faktorer.
Många joniska kanaler och transportproteiner styrs direkt av proteinproteiner G. På liknande sätt är dessa proteiner involverade i många sensoriska processer som syn, lukt, bland andra.
Hur fungerar de?
Läget för interaktion mellan ett G -protein med effektorproteiner är typiskt för varje klass eller proteinfamilj.
För G-grebranala receptor G-proteiner (heterotrofimeriska G-proteiner) orsakar föreningen av en guaninknleotid såsom BNP eller Guanosín-difosfat till underenheter a förening av de tre underenheterna, som bildar ett komplex som kallas GαPy eller G-GNP, som är länkad till membranet.
Om BNP-molekylen därefter utbyts av en GTP-molekyl, är a-underenheten ansluten till GTP-dissociat från p- och y-underenheterna, och bildar ett separat komplex som kallas Gα-GTP, som kan förändra aktiviteten hos dess enzymer eller Diana-transportband proteiner.
Den hydrolytiska aktiviteten för denna underenhet gör att du kan avsluta aktiveringen, utbyta GTP för en ny BNP, flytta till den inaktiva konformationen.
I avsaknad av de upphetsade receptorerna som är associerade med BNP: s BNP -utbytesprocess är det mycket långsamt, vilket innebär att heterotrimiska GD -proteiner bara utbyter BNP med GTP med en fysiologiskt viktig hastighet när de förenas med sina upphetsade receptorer.
Referenser
- Gilman, G. (1987). G-proteiner: överföringar av mottagar-genererade signaler. Årliga recensioner i biokemi, 56, 615-649.
- Milligan, g., & Kostenis, och. (2006). Heterotrimiska G-proteiner: En kort historia. British Journal of Pharmacology, 147, 546-555.
- Offermanns, s. (2003). G-proteiner som överföringar i transmembransignalering. Framsteg inom biofysik och molekylärbiologi, 83, 101-130.
- Simon, m., Strathmann, m. P., & Gautam, n. (1991). Mångfald av G -proteiner i signaltransduktion. Vetenskap, 252, 802-808.
- Syrovatkina, v., Glada, k. ANTINGEN., Dey, r., & Huang, x. (2016). Reglerings-, signal- och fysiologiska funktioner hos G-proteiner. Journal of Molecular Biology, 428(19), 3850-3868.
- « Crotalus triseriatusegenskaper, livsmiljö, reproduktion
- Åska egenskaper, livsmiljö, taxonomi, egenskaper »