

DNA -replikationsmekanismer, i prokaryoter och eukaryoter
- 4378
- 626
- PhD. Emil Svensson
De Replikering av DNA (Desoxyribonukleinsyra) Den består av att kopiera genomet, det vill säga all genetisk information som finns i DNA från en organisme, för att producera två identiska kopior. Genomet har nödvändig information för att bygga en komplett organisme.
Innan celldelning inträffar DNA -replikering. Genom meios inträffar gameter för sexuell reproduktion. Genom mitos sker cellersättning (till exempel hud och blod) och utveckling (till exempel vävnader och organ).
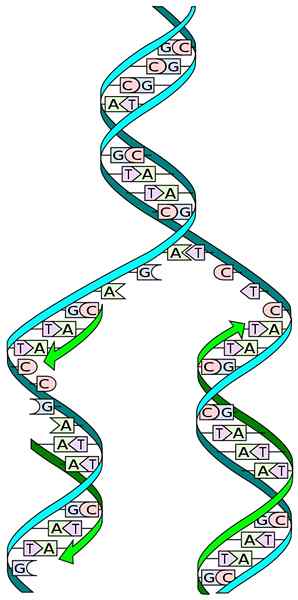
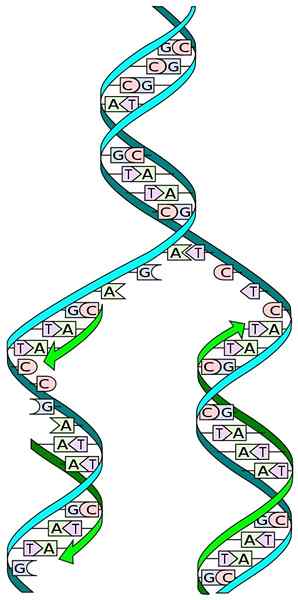 Källa: I, Madprime [CC0]
Källa: I, Madprime [CC0] Kunskapen om DNA -strukturen gör att vi kan förstå hur dess replikering inträffar. Strukturen för DNA består av en dubbel propell, sammansatt av två antipaallala kedjor av successiva nukleotider, vars kvävebaser kompletteras specifikt.
Under replikationen fungerar var och en av kedjorna i den dubbla DNA -kedjan som en form för biosyntesen av en ny kedja. De två nyligen syntetiserade kedjorna har baser som är komplementära till baserna i mögelkedjan: adenin (A) med Timina (T) och cytosin (C) med guanin (G).
Flera enzymer och proteiner deltar i DNA -replikation. Att till exempel öppna den dubbla DNA-propellen, hålla DNA öppet och tillsätta deoxyribonukleosidos-5'-trifosfat (DNTP) för att bilda den nya kedjan.
[TOC]
DNA -replikering är semi -konservativ
Baserat på DNA: s struktur föreslog Watson och Crick att DNA -replikering sker på ett semi -konservativt sätt. Detta demonstrerades av Meselson och Stahl genom att markera DNA från Escherichia coli Med den tunga isotopen av kväve, femtonN, följer i flera generationer distributionsmönstret i ett odlingsmedium med lätt kväve, 14N.
Messelson och Stahl fann att i den första generationen hade de två döttrarna DNA -molekyler, varje molekyl markerad med en kedja med den tunga kväveisotopen och en annan med den lätta isotopen. Till skillnad från föräldradna -molekylen, som hade de två kedjorna markerade med den tunga isotopen, femtonN.
I den andra generationen var 50% av DNA -molekylerna som de i den första generationen, och de andra 50% hade bara lätt kväve. Tolkningen av detta resultat är att den dubbla dotterpropellen har en föräldrakedja (som fungerar som en form) och en ny kedja.
Den semi -konservativa replikationsmekanismen innebär separering av DNA -kedjor och parning av kompletterande baser med hjälp av successiva nukleotider, vilket producerar två dubbla döttrar döttrar döttrar.
Replikering i batterier
Start av DNA -replikering i bakterier
Bakterier DNA består av en cirkulär kromosom och har bara en plats för replikationsstället. Från denna webbplats inträffar biosyntesen av de två döttrarna dubbelriktat och bildar två replikationsgafflar som rör sig i riktningar i motsats till ursprunget. I slutändan är gafflarna att slutföra replikationen.
Replikering börjar med föreningen av DNAA -proteiner till ursprungsplatsen. Dessa proteiner bildar i sin tur ett komplex. Då binder Hu- och IHF -proteinerna.
Därefter förenas DNAC -proteiner, vilket gör att Helicases of DNA går med. Dessa hjälper till att varna av DNA och bryta vätebindningarna, bildade i baspar. Så de två kedjorna separerar ännu mer och bildar två enkla kedjor.
Kan tjäna dig: vad är kromosomal permutation?Topoisomeras II, eller DNA Girasa, rör sig framför DNA Helicy. SSB -proteiner (enkel kedja DNA -bindning (SSB) håller DNA -kedjorna åtskilda. Således kan biosyntesen av dotterkedjan börja.
Biosyntes av döttrar DNA -kedjor i bakterier
Prima -enzym är ansvarigt för att syntetisera korta RNA -kedjor som kallas primrar, som har 10 till 15 nukleotider i längd. DNA-polymeras börjar lägga till 5'-trifosfat (DNTP) till 3'-OH för primningssockret, varefter kedjan fortsätter att växa vid samma ändamål.
Eftersom DNA -kedjor är antiparallella, syntetiseras en primer i guidekedjan och många primrar i den försenade kedjan. På grund av detta är biosyntesen av den försenade kedjan diskontinuerlig. Även om DNA -kedjor är antiparallella, rör sig replikationsgaffeln i en riktning.
DNA -polymeraset är ansvarigt för bildandet av kovalenta bindningar mellan angränsande nukleotider av de nyligen syntetiserade kedjorna, i 5'®3 'riktning. I OCH. coli, Det finns fem polymeras -DNA: DNA -polymeraser I och III utför DNA -replikation; och DNA -polymeraserna II, IV och V är ansvariga för att replikera och replikera skadat DNA.
Det mesta av replikationen tillverkas av DNA -polymeras III, som är ett holoenzym som har 10 olika underenheter med flera funktioner i replikationen av DNA. Till exempel är Alpha -underenheten ansvarig för att göra bindningar mellan nukleotider.
Ett enzymkomplex ansvarar för replikering av DNA i bakterier
Helikaset av DNA och Prima förenas för att bilda ett komplex som kallas Primosoma. Detta rör sig längs DNA och agerar på ett samordnat sätt att separera de två föräldrakedjorna och syntetisera primrarna varje viss intervall på den försenade kedjan.
Primosom binder fysiskt till DNA -polymeras III och bildar det replisome. Två DNA -polymeraser III är ansvariga för att replikera DNA för guiden och försenade kedjor. Med avseende på DNA -polymeras III bildar den försenade kedjan en slinga, vilket gör att tillägg av nukleotider till denna kedja kan hända i samma riktning för styrkedjan.
Tillsatsen av nukleotider till styrkedjan är kontinuerlig. Medan det är i förseningen är det diskontinuerligt. Fragment av 150 nukleotider av längd bildas, kallade Okazaki -fragment.
Exonukleasaktiviteten 5 ' -> 3' av DNA -polymeras I är ansvarig för att eliminera primrarna och fylla, tillsätta nukleotider. Ett enzymligas tätar luckorna mellan fragment. Replikering slutar när de två replikeringshoquillorna är i en färdigställningssekvens.
Proteinet som du binder till avslutningssekvensen och stoppar förflyttningen av replikationsgaffeln. Topoisomeras II tillåter separering av de två kromosomerna.
Desoxiribonukleotider Tryfosfater används av DNA -polymeras
Dexinukleosid tryfosfat (DNTP) innehåller tre fosfatgrupper förenade till kol 5 'av deoxyrbosa. DNTPS (DATP, DTTP, DGTP och DCTP) går med i mögelkedjan efter AT/GC -regeln.
Kan tjäna dig: plesiomorphyDNA-polymeraset katalyserar följande reaktion: hydroxylgruppen (-OH) 3 'av nukleotiden i den växande kedjan reagerar med alfa-fosfatet i det inkommande DNTP, frisläppande oorganiskt pyrofosfat (PPI) (PPI). PPI -hydrolys producerar energi för bildandet av den kovalenta bindningen, eller fosfodiésterbindning, mellan nukleotider i den växande kedjan.
Mekanismer som säkerställer replikationens trohet hos DNA
Under DNA -replikering gör DNA -polymeras III ett fel per 100 miljoner nukleotider. Även om sannolikheten för fel är mycket låg, finns det mekanismer som säkerställer trohet i DNA -replikation. Dessa mekanismer är:
1) Stabilitet i bas Apar. Vätebindningsenergin mellan AT/GC är större än i felaktiga baspar.
2) DNA -polymerasaktiv platsstruktur. DNA -polymeras katalyserar företrädesvis nukleotider med korrekta baser i motsatt kedja. En dålig parning av baser orsakar en snedvridning av den dubbla DNA -propellen, vilket förhindrar att den felaktiga nukleotiden ockuperar det aktiva stället för enzymet.
3) Läsningstest. DNA -polymeras identifierar införlivade felaktiga nukleotider och eliminerar dem från dotterkedjan. Exonukleasaktiviteten för polymeras -DNA bryter fosfodiésterbindningarna mellan nukleotider vid 3' -änden av den nya kedjan.
DNA -replikering i eukaryoter
Till skillnad från replikationen i prokaryoter, vars replikering börjar på en enda plats, börjar eukaryotisk replikering på flera ursprungsplatser och replikeringsgaffeln rör sig dubbelriktad. Därefter slås alla replikationsgafflar sammanslagna och bildar två systerkromatider som förenats i centromeren.
Eukaryotas har många typer av DNA -polymeras, vars namn använder grekiska bokstäver. DNA -polymeras a bilda ett komplex med prima. Denna komplexa syntetiserar korta primrar bestående av 10 RNA -nukleotider följt av 20 till 30 DNA -nukleotider.
Sedan DNA -polymeras ε antingen Δ katalyserar förlängningen av dotterkedjan från primern. DNA -polymeras ε Det är involverat i syntesen av den ledande kedjan, medan DNA -polymeras Δ Syntetisera den försenade kedjan.
DNA -polymeras Δ Förlänger vänster Okazaki -fragment tills den når den högra RNA -primern, vilket ger en kort lyftning av primern ("Kort klaff"). Till skillnad från prokaryoter, där ett polymeras -DNA eliminerar primern, eliminerar i eukaryoterna en endonukleasflikenzym.
Därefter tätar en DNA -ligas DNA -fragmenten som är intilliggande. Avslutningen av replikering sker med dissociation av replikationsgaffelproteiner.
De DNA -replikering i eukaryoter och cellcykel
Replikering i eukaryoter sker i S -fasen av cellcykeln. Replikerade DNA -molekyler är segregerade i två dotterceller under mitos. Faser G1 och G2 separerar S -fasen och mitosen. Progressionen genom varje fas i cellcykeln är starkt reglerad av kinaser, fosfataser och proteaser.
I G1 -fasen i cellcykeln ansluter sig ursprungsigenkänningskomplexet (OCR) till ursprungsplatsen. Detta inducerar Union of MCM Helicas och andra proteiner, såsom CDC6 och CDT1, för att bilda ett förreplikationskomplex (PRERC). Las Helicase MCM går med i guidekedjan.
Kan tjäna dig: länkade generI fas S blir PRERC en aktiv replikationsplats. OCR-, CDC6- och CDT1 -proteiner frisätts och MCM -helikaset rör sig i riktning av 3 'till 5'. När replikationen är slut kommer detta att startas om i nästa cellcykel.
Replikering av ändarna på kromosomer i eukaryoter
Ändarna på kromosomerna är kända som telomerer, som består av upprepade sekvenser i tandem och av en 3' -region som utmärker sig, från 12 till 16 nukleotider av längd.
DNA -polymeras kan inte replikera 3' -slutet av DNA -kedjor. Detta beror på att DNA-polymeras endast kan syntetisera DNA i adressen 5'-3 'och endast kan förlänga befintliga kedjor, utan att kunna syntetisera en primer i denna region. Följaktligen förkortas telomerer i varje replikationsrunda.
Telomerasenzym förhindrar telomerer förkortning. Telomeras är ett enzym som har protein- och RNA -underenheter (tredje). Den senare binder till de upprepade sekvenserna av DNA och gör att telomeras kan sammanfogas till 3' -änden av telomeren.
En sekvens av RNA bakom unionen belägen som en form för syntesen av en sekvens av sex nukleotider (polymerisation) i slutet av DNA -kedjan. Förlängningen av telomeren katalyseras av underenheter av telomeras, kallad omvänd telomerastranskriptas (TERT).
Efter polymerisation sker translokalisering, bestående av rörelsen av telomeras till en ny ände av DNA -kedjan och förenar sig med sex andra nukleotider fram till slutet.
Funktionerna för andra DNA -polymeraser i eukaryoterna
DNA -polymeras p Det har en viktig roll i eliminering av felaktiga baser av DNA, men det är inte involverat i DNA -replikering.
Många upptäckte polymeras-DNA tillhör den "translesionsreplikerande" polymerasgruppen. Dessa polymeraser ansvarar för att syntetisera kompletterande kedjor i ett skadat DNA -region.
Det finns flera typer av polymeraser "translesion-replikerande". Till exempel DNA -polymeras η Du kan replikera om Timina Dímeros, som produceras av UV Light.
DNA -replikering i arkeobakterier
Replikationen av arkeobakteria -DNA liknar den som ges i eukaryoter. Detta beror på följande: 1) proteiner som deltar i replikering är mer som de hos eukaryoter än på prokaryoter; och 2) Även om det bara finns en replikationsplats som i prokaryoter, liknar dess sekvens platsen för eukaryot ursprung.
Likheten i replikationen mellan bågarna och eukaryoterna stöder idén att båda grupperna är fylogenetiskt mer relaterade till varandra än någon av dem med prokaryoterna.
Referenser
- Brooker, r. J. 2018. Genetikanalys och principer. McGraw-Hill, New York.
- Hartwell, L. H., Goldberg, M. L., Fischer, J. TILL., Hood, L. 2018. Genetik - från genomgener. McGraw-Hill, New York.
- Kušić-Tišma, J. 2011. Grundläggande aspekter av DNA -replikation. Intech öppen åtkomst, Kroatien.
- Lewis, r., 2015. Mänskliga genetikbegrepp och tillämpningar. McGraw-Hill, New York.
- Pierce, b. TILL. 2005. Genetik - En konceptuell strategi. W. H. Freeman, New York.