

Ribulosa -egenskaper, struktur och funktioner
- 1619
- 96
- Johan Olsson
De Ribulosa Det är ett monosackaridsocker eller kolhydrat som innehåller fem kolatomer och en ketonfunktionell grupp i dess struktur, så den ingår i Ketopenticas -gruppen.
Kyrkan med fyra och fem kolatomer namnges genom att infoga infixen "Ul”I namnet motsvarande Aldosa. Så, d-libulous är ketopentosa som motsvarar d-liboset, en aldopentosa.
Fisher -projektion för ribulosa (källa: Neurotoger [Public Domain] via Wikimedia Commons) Detta socker deltar i D-libulous formen som en mellanhand i olika metaboliska rutter, som i Calvin-cykeln, till exempel. Medan ensam i vissa genrer bakterier som Acetobacter och Glukonobacter L-libos erhålls som en slutlig metabolisk produkt. Av denna anledning används dessa mikroorganismer för sin syntes på industriell nivå.
Vissa föreningar härrörande från ribulosen är av de viktigaste mellanhandsföreningarna på pentosfosfatens väg. Denna rutt är avsedd att generera NADPH, en viktig kofaktor som fungerar i nukleotidbiosyntes.
Det finns industriella mekanismer för att syntetisera l-libulous som isolerad förening. Den första isoleringsmetoden som den erhölls bestod av Levene- och Forge Zeal-isoleringsmetoden från L-Xilosa.
Trots de stora framstegen i industriella metoder för syntes och rening av kemiska föreningar erhålls inte L-libulous som isolerad monosackarid, vilket erhålls i kombinerade fraktioner av L-libos och L-yrabinous.
Metoden för att få den mest använda libulous idag är rening från gLuconobakt Frarateurii IFO 3254. Denna typ av bakterier kan överleva under sura förhållanden och ha en ribitoloxidationsväg till L-libulous.
Det kan tjäna dig: sinaloa flora och fauna: vanligare djur och växter[TOC]
Egenskaper
Ribulous Som ett syntetiserat, extraherat och renat reagens som ofta finns som l-libulous, är ett solid, vit och kristallin organisk substans. Liksom alla kolhydrater är denna monosackarid vattenlöslig och har de typiska egenskaperna hos polära ämnen.
Som är vanligt för resten av sackariderna har ribulosen samma antal kol- och syreatomer, och två gånger denna mängd i väteatomer.
Det vanligaste sättet på vilket det finns i ribulösa i naturen är i samband med olika substituenter och bildar komplexa strukturer, vanligtvis fosforylerade, såsom ribulösa 5-fosfat, ribulösa 1,5-biskfosfat, bland andra.
Dessa föreningar fungerar vanligtvis som mellanhänder och transportörer eller "fordon" för fosfatgrupper i de olika cellulära metaboliska vägarna där de deltar.
Strukturera
Ribulosa-molekylen har ett centralt skelett av fem kolatomer och en cetona-grupp i kol i position C-2. Som tidigare nämnts placerar denna funktionella grupp den inom ketosas som ketopentosa.
Den har fyra hydroxylgrupper (-OH) förenade till de fyra kol som inte är fästa vid ketongruppen och dessa fyra kol är mättade med väteatomer.
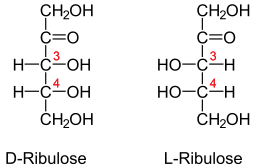
Den ribulösa molekylen kan representeras enligt projektionen av Fisher i två former: D-libulous eller libulous, formen är stereoisomer och enantiomer i formen D och vice versa.
Klassificeringen av D- eller L -formen beror på orienteringen av hydroxylgrupperna i den första kolatomen efter Cetona -gruppen. Om denna grupp är inriktad på höger sida, motsvarar molekylen på Fisher-uppdraget D-libulous, annars om den ligger till vänster (L-libulous).
Det kan tjäna dig: Vilka är skillnaderna mellan fotosyntes och andning?I Haworths projektion kan ribulosa representeras i ytterligare två strukturer beroende på orientering av hydroxylen i den anomera kolatomen. I p -positionen är hydroxylen orienterad mot den övre delen av molekylen; Medan a -positionen styr hydroxyl mot botten.
Enligt Haworth-projektionen kan således fyra möjliga sätt tas: ß-d-libulous, a-d-libulous, ß-l-libulosa eller a-l-libulous.
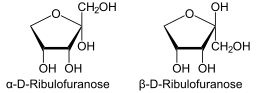 Haworth -projektion för Ribulofuranosa (källa: Neurotokeker [Public Domain] via Wikimedia Commons)
Haworth -projektion för Ribulofuranosa (källa: Neurotokeker [Public Domain] via Wikimedia Commons) Funktioner
Pentosfosfatväg
De flesta av cellerna, särskilt de som finns i snabb och konstant uppdelning, såsom benmärg, tarmslemhinnor och tumörceller, använder ribulös-5-fosfat, som isomeriseras till ribos-5-fosfat i oxidativ väg för pentosfosfat , för att producera nukleinsyror (RNA och DNA) och koenzymer såsom ATP, NADH, FADH2 och Coenzima A.
Denna oxidativa fas av pentosfosfat innehåller två oxidationer som omvandlar 6-fosfatglukos till ribulosa 5-fosfat, vilket reducerar NADP+ till NADPH.
Dessutom är ribulosa-5-fosfat aktivt indirekt.
Calvin -cykel
Calvin -cykeln är kolfixeringscykeln som äger rum i fotosyntetiska organismer efter de första reaktionerna av fotosyntes.
Det har bevisats genom att markera metoder i studier som genomförts av olika forskare, som genom att markera kol i C-1-positionen för ribulosa-1,5-bifosfat, är koldioxiden inställd i denna mellanhand under Calvin-cykeln till två Molekyler av 3-fosfoglycerate: en märkt och en utan att markera.
Rubisco (ribulosa 1,5-biskoposfatkarboxylas/syretas) anses vara det vanligaste enzymet på planeten och använder som ett substrat det ribulösa 1,5-biskosfatet, för att katalysera införlivandet av koldioxid och produktion av 1,3-diffoglykerat i Calvincykeln.
Det kan tjäna dig: Lipase: Caracateristisk, struktur, typer, funktionerBrottet av denna instabila mellanhand, 1,3-diffoglycerat, av sex kolatomer, katalyseras också av Rubisco, som är den som förmedlar bildningen av två molekyler av 3 kolatomer (3-fosfoglycerat).
Funktioner i bakterier
Enol-1-ANTINGEN-Karboxifenyl. I detta steg släpps en koldioxidmolekyl och en av vatten, och producerar också en Indol-3-glycerol-fosfatmolekyl.
Bakterier använder l-libulous också på de rutter som används för etanolmetabolism. Dessutom har dessa mikroorganismer ett enzym som kallas L-arabinöst isomeras, som modifierar arabinalt för att syntetisera l-libulous.
L-libulosa quinasa fosfororila Denna metabolit nedströms för att bilda L-5-fosfat, som kan komma in i vägen för pentosfosfat för produktion av sockerarter för skelett av nukleinsyror och andra väsentliga molekyler.
Referenser
- Ahmed, Z. (2001). Produktion av naturliga och sällsynta pontoser med hjälp av mikroorganismer och deras enzymer. Elektronisk tidskrift för bioteknik, 4(2), 13-14.
- Ahmed, Z., Shimonishi, T., Buiyan, s. H., Utamura, m., Takada, g., & Izumori, k. (1999). Biokemisk beredning av l-libos och l-yarabinose från ribitol: en ny strategi. Journal of Bioscience and Bioengineering, 88(4), 444-448
- Finch, s. (Ed.). (2013). Kolhydrater: strukturer, syntesser och dynamik. Springer Science & Business Media.
- Murray, r., Bender, D., Botham, K. M., Kennelly, s. J., Rodwell, V., & Weil, s. TILL. (2012). Harpers Illustrated Biochemistry 29/E. Ed Mc Graw Hill Lange, Kina
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger principer för biokemi. Macmillan.
- Stick, r. V. (2001). Kolhydraater: The Sweet Molecules of Life. Annars.