

Syntes av lipidtyper och deras huvudmekanismer
- 1186
- 89
- Lars Eriksson
De Lipidsyntes Den består av en serie enzymatiska reaktioner med vilka kortkedjiga kolväten kondenseras för att bilda längre kedjemolekyler som därefter kan drabbas av olika kemiska modifieringar.
Lipider är en mycket varierad typ av biomolekyler syntetiserade av alla levande celler och är specialiserade på flera väsentliga funktioner för att upprätthålla celllivet.
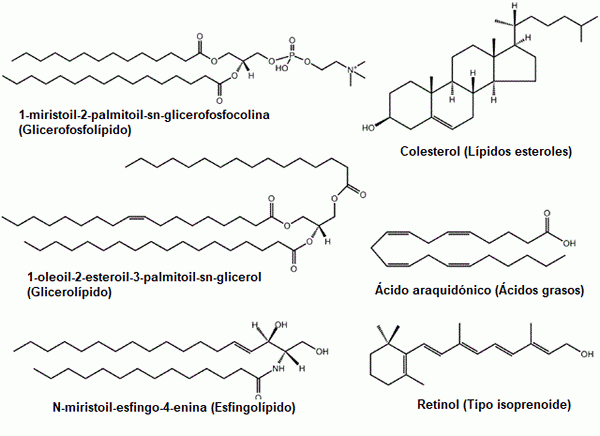 Några exempel på vanliga lipider: glycerofosfolipider, steroler, glycerolipider, fettsyror, sfingolipider och prenoler (källa: den ursprungliga uppladdaren var lmaps på engelska wikipedia. / Gfdl 1.2 (http: // www.gnu.Org/licenser/gamla licenser/FDL-1.2.html) via Commons, anpassad av Raquel Parada)
Några exempel på vanliga lipider: glycerofosfolipider, steroler, glycerolipider, fettsyror, sfingolipider och prenoler (källa: den ursprungliga uppladdaren var lmaps på engelska wikipedia. / Gfdl 1.2 (http: // www.gnu.Org/licenser/gamla licenser/FDL-1.2.html) via Commons, anpassad av Raquel Parada) Lipider är de viktigaste komponenterna i biologiska membran, ett faktum som gör dem till grundläggande molekyler för förekomsten av celler som isolerade enheter i deras omgivningar.
Vissa lipider har också specialiserade funktioner som pigment, kofaktorer, transportörer, tvättmedel, hormoner, intra-extrakellulära budbärare, kovalenta ankare för membranproteiner etc. Därför är förmågan att syntetisera olika typer av lipider avgörande för överlevnaden för alla levande organismer.
Denna stora grupp av föreningar klassificeras traditionellt i flera kategorier eller undergrupper: fettsyror (mättade och omättade), glyceridkomplex lipider (lipoproteiner).
[TOC]
Typer av lipider och deras huvudsakliga syntesmekanismer
Alla reaktionssekvenser av lipidbiosyntesvägar är endogoniska och reduktiva. Med andra ord, de använder alla ATP som en energikälla och en reducerad elektrontransportör, såsom NADPH, som en reducerande kraft.
Därefter beskrivs de viktigaste reaktionerna i de biosyntetiska vägarna för huvudtyperna av lipider, det vill säga av fett- och eikosanoidsyror, triacylglyceroler och fosfolipider och steroler (kolesterol) (kolesterol).
- Fettsyrasyntes
Fettsyror är oerhört viktiga molekyler från lipidsynpunkten, eftersom de är en del av de mest relevanta lipider i celler. Hans syntes, i motsats till vad många forskare trodde under de första studierna, består inte av den omvända vägen för deras ß-oxidation.
I själva verket inträffar denna metaboliska väg i olika cellfack och kräver deltagande av en mellanhand av tre kolatomer som kallas malonyl-CoA, vilket inte är nödvändigt vid oxidation.
 Malonylkoa. Neuroteker / pubdomän
Malonylkoa. Neuroteker / pubdomän Dessutom är det nära besläktat med sulfhydrilgrupperna av proteiner kända som acylgruppstransportörer (ACP, engelska Acylbärarproteiner).
I allmänhet är syntesen av fettsyror, särskilt den för den långa kedjan, en sekventiell process där fyra steg upprepas i varje "återvändande", och under varje återkomst finns en mättad syrogrupp som är underlaget för nästa, som innebär en annan kondensation med en ny malonyl-CoA-molekyl.
I varje sväng eller reaktionscykel sträcker sig fettsyrakedjan två kol, tills en längd på 16 atomer (palmitate) når, varefter cykeln lämnar.
Malonyl-CoA-bildning
Denna mellanhand av tre kolatomer är irreversibelt bildad av acetyl-CoA tack vare verkan av en acetyl-CoA-karboxylasenzym, som har en biotingrostatgrupp som är kovalent kopplad till enzymet och som deltar i denna katalys i denna katalys i två steg.
I denna reaktion överförs en karboxylgrupp härledd från en bikarbonatmolekyl (HCO3-) till biotinet vid en ATP-beroende form, där biotinylgruppen uppfyller funktionen "tillfällig transportör" av molekylen medan överföringar till acetyl-CoA, producerar malonyl-CoA.
Kan tjäna dig: oxidastest: grund, procedur och användningarI sekvensen av syntes av fettsyror är det reducerande medel som används NADPH och de aktiverande grupperna är två tiolgrupper (-sh) som är en del av ett multienzimatiskt komplex som kallas syntasfettsyra, som är det som är viktigast i katalys syntetik.
I ryggradsdjur är syntasfettsyrakomplexet en del av en enda stor polypeptidkedja, där de 7 enzymatiska aktiviteterna som är karakteristiska för syntesvägen representeras, liksom den hydrolytiska aktivitet som krävs för att frigöra mellanhänder i slutet av syntesen.
 Syntasa-fettsyranzymstruktur (källa: Boehringer Ingelheim/CC BY-SA (https: // Creativecommons.Org/licenser/BY-SA/4.0) via Wikimedia Commons)
Syntasa-fettsyranzymstruktur (källa: Boehringer Ingelheim/CC BY-SA (https: // Creativecommons.Org/licenser/BY-SA/4.0) via Wikimedia Commons) De 7 enzymatiska aktiviteterna för detta komplex är: ACILO-grupptransportprotein (ACP), acetyl-CoA-ACP transacetilas cetoacil-ACP-reduktas (KR), ß-hydroxyacil-ACP dehydratasa (HD) och ilska-ACP-reduktas (ER).
Innan kondensationsreaktionerna kan inträffa för att montera fettsyrakedjan överförs de två tiolgrupperna i det enzymatiska komplexet med acylgrupperna: först och främst överförs en acetyl -COA till -sh -gruppen av en cystein I p-cetoacyl-ACP-syntasen av komplexet katalyserat reaktion katalyserat av acetyl-CoA-ACP-transacetylaenzymet (AT) (AT).
Därefter överförs en malonisk grupp från en malonyl -CoA -molekyl till -sh -gruppen i ACILO -grupptransportören är en del av fettsyrakomplexet syntas.
Sekvensen av fyra reaktioner för varje "retur" av reaktionscykeln är som följer:
- Kondensation: Acetyl- och malonylgrupper "laddade" i enzymet kondenseras för att bilda en acetoacetyl -ACP -molekyl, som är kopplad till ACP -delen genom en grupp -sh. I detta steg finns en CO2-molekyl och katalyseras av ß-Zoacyl-ACP-syntasen (Acetylgruppen upptar positionen som ”terminal metyl” i acetoacetyl-ACP-komplexet)).
- Reduktion av karbonylgruppen: Karbonylgruppen i C3-positionen för acetoacetyl-ACP reduceras till bildning av D-p-hydroxibutiril-ACP, reaktion katalyserad av p-cetoacyl-ACP-reduktaset, som NADPH använder som elektron donor.
- Dehydrering: C2- och C3-kol i D-p-hydroxibutiril-ACP saknar vattenmolekyler och bildar en dubbelbindning som slutar med produktionen av den nya föreningen trans-∆2-butenoil-ACP. Denna process medieras av ett p-hydroxyacyl-ACP dehydratas (HD) enzym (HD).
- Dubbelbindningsreduktion: Den dubbla bindningen för föreningen som bildas i dehydreringssteget är mättat (reducerat) för att ge upphov till butiril-ACP av reaktionen katalyserad av enzymet-ACP-reduktaset (ER), som också använder NADPH som reducerande medel.
Syntesreaktionerna inträffar tills en palmitatmolekyl (16 kolatomer) bildas, som är hydrolyserad av det enzymatiska komplexet och frigörs som en möjlig föregångare för fettsyror med större längdkedjor, som produceras av töjningssystem av fettsyror belägna i smidigt del av endoplasmatisk retikulum och i mitokondrierna.
Det kan tjäna dig: fauna och flora i det peruanska havetDe andra modifieringarna som dessa molekyler kan drabbas av till exempel deaturationerna, katalyseras till exempel av olika enzymer, som i allmänhet förekommer i den släta endoplasmiska retikulumen.
- Syntes av eikosanoider
Eikosanoider är celllipider som har funktioner som "kort -range" messenger -molekyler, producerade av vissa tyger för att kommunicera med sina angränsande tyger. Dessa molekyler syntetiseras från fleromättade fettsyror med 20 kolatomer.
Prostaglandiner
Som svar på en hormonell stimulans attackerar fosfolipasenzymet membranfosfolipider och frisätter araquidonato från Glycerol Carbon 2. Denna förening omvandlas till prostaglandiner tack vare ett enzym av den släta endoplasmiska retikulum med bifunktionell aktivitet: cyklooxygenas (COX) eller prostaglandin H2 -syntas.
Tromboxan
Prostaglandins kan omvandlas till tromboxaner tack vare tromboxansyntas i blodplättarna (trombocyter). Dessa molekyler deltar i de första stegen för blodkoagulering.
- Syntes av triacylglyceroler
Fettsyror är grundläggande molekyler för syntes av andra mer komplexa föreningar i celler, såsom triacylglyceroler eller lipidglycerofosfolipidlipidermembran (processer som beror på cellulära metaboliska behov).
Djur producerar triacylglyceroler och glyceofosfalipider från två vanliga föregångare: fett-CoA-acyl och 3-fosfat L-glycerol. Fat-CoA-acyl produceras av acyl-CoA-synteser som deltar i ß-oxidation, medan 3-fosfat L-glycerol erhålls från glykolys och genom verkan av två alternativa enzymer: 3-fosfatglyceroldehydrogenas och kinasglykcerol.
Tiacylglycerolerna bildas av reaktionen mellan två molekyler av flo-CoA-acil och en 3-fosfatdiacylglycerolmolekyl; Dessa överföringsreaktioner katalyseras av specifika transferas.
I denna reaktion finns det initialt fosfatidinsyra, som defosforyleras av ett enzymfosfatidsyrafosfatas för att producera 1,2-diacylglycerol, som återigen kan acceptera en tredje molekyl av fett-CoA-acyl, vilket producerar triacilglyceroll.
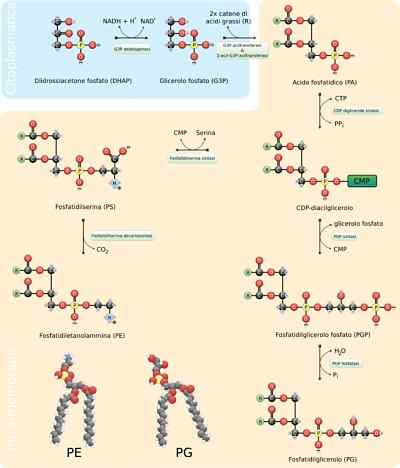
- Syntes av fosfolipider
Fosfolipider är extremt varierande molekyler, eftersom många olika kan bildas genom kombinationen av fettsyror och "huvud" -grupper med glycerolskelettet (glyceofosfolipider) eller sfinosin (sfingolipider) som kännetecknar dem.
Generalförsamlingen för dessa molekyler kräver syntes av skelettet av glycerol eller sfärisk, av föreningen med motsvarande fettsyror, antingen genom sterifiering eller amidering, tillägg av en hydrofil "huvud" -grupp genom en fosfodiésterbindning och, om nödvändigt , förändring eller utbyte av dessa sista grupper.
I eukaryoter sker denna process i den släta endoplasmiska retikulum och även i det inre mitokondriella membranet, där de kan förbli på obestämd tid eller där de kan omlokeras till andra platser.
Reaktionssteg
De första stegen i syntesreaktionen av glycerofosfolipider är ekvivalent med de för triacylglycerolproduktion, eftersom en 3-fosfatglycerolmolekyl förstörs till två fettsyramolekyler i kol 1 och 2, vilket bildar fosfatidinsyran. Det är vanligt att hitta fosfolipider som har mättade fettsyror i C1 och omättas i C2 av glycerol.
Kan tjäna dig: resistinaFosfatidinsyra kan också produceras genom fosforylering av en diacylglycerolmolekyl som redan syntetiseras eller "återvinns".
"Huvud" polära grupper av dessa molekyler bildas genom fosfodiésterlänkar. Det första som borde hända för att denna process ska vara korrekt är "aktiveringen" av en av de hydroxylgrupper som deltar i processen med hjälp av hydroxyl som deltar i reaktionen.
Om denna molekyl binder till diacylglycerol bildas CDP-diacylglycerol (den "aktiverade" formen av fosfatidinsyra), men detta kan också hända på hydroxylgruppen i gruppen "huvud".
I fallet med fosfatidylserin aktiveras till exempel fördröjningscerol genom kondensation av fosfatidinsyramolekylen med en trif-citidincytidinmolekyl (CTP), bildar CDP-diacylglycerol och eliminerar en pyrofosfat.
Om en MMP-molekyl (cytidinmonofosfat) rör sig genom en nukleofil attack av serinen eller hydroxylhydroxylen i kol 1 i 3-fosfatglycerolen. Fosfat producerar fosfatidylglycerol.
Båda molekylerna som produceras på detta sätt fungerar som föregångare för andra membranlipider, som ofta delar biosyntetiska vägar från varandra.
- Kolesterolsyntes
Kolesterol är en väsentlig molekyl för djur som kan syntetiseras av dess celler, så det är inte viktigt i den dagliga dieten. Denna molekyl med 27 kolatomer produceras från en föregångare: acetat.
Denna komplexa molekyl bildas av acetyl-CoA i fyra huvudstadier:
- Kondensation av tre acetatenheter för att bilda mevalonato, en mellanhandsmolekyl av 6 kol (först bildas en acetoacetyl-CoA-molekyl med två acetyl-CoA (tiolasa enzym) och sedan en annan av p-hydroxi-p-metylglutaril-CoA (HMG- COA) (HMG-CoA-synteta enzym). Mevalonato bildas från HMG-CoA och tack vare HMG-CoA-reduktasenzymet.
- Mevalonato -omvandling till isoprenenheter. Första 3 fosfatgrupper överförs från 3 ATP -molekyler till Mevalonato. En av fosfaterna går förlorad tillsammans med den angränsande karbonylgruppen och bildas
- Polymerisation eller kondensation av 6 isoprenenheter av 5 kolatomer för att bilda 30 -kolatomer (en linjär molekyl) rakning (en linjär molekyl).
- Cicration av escualean för att bilda de fyra ringarna i steroidkärnan i kolesterol och efterföljande kemiska förändringar: oxidationer, migration och eliminering av metylgrupper etc., Vad som ger kolesterol.
Referenser
- Garrett, R. H., & Grisham, c. M. (2001). Principer för biokemi: med ett mänskligt fokus. Brooks/Cole Publishing Company.
- Murray, r. K., Granner, D. K., Mayes, s. TILL., & Rodwell, V. W. (2014). Harpers illustrerade biokemi. McGraw-hill.
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger principer för biokemi. Macmillan.
- Jacquemyn, J., Cascaho, a., & Goodchild, r. OCH. (2017). INS och outs av endoplasmatisk retikulum - kontrollerad lipidbiosyntes. Embo Reports, 18 (11), 1905-1921.
- Ohlrogge, j., & Bläddra, J. (nittonhundranittiofem). Lipidbiosyntes. Växtcellen, 7 (7), 957.
- « José López Portillo Biografi, regering och bidrag
- Tropisk djungel i Mexiko egenskaper, plats, lättnad, flora, fauna »