

Ketogenes typer av kroppar, syntes och nedbrytning
- 3267
- 972
- Karl Johansson
De ketogenes Det är processen genom vilken acetoacetat, p-hydroxibutirat och aceton erhålls, som tillsammans kallas ketonkroppar. Denna komplexa och fint reglerade mekanism utförs i mitokondrier, från katabolismen av fettsyror.
Att få ketonkropparna äger rum när kroppen utsätts för uttömmande fasta perioder. Även om dessa metaboliter mestadels syntetiseras i leverceller, finns de som en viktig källa till energikälla i olika vävnader, såsom skelettmuskler och i hjärta och cerebrala vävnader.
Källa: Sav vas [CC0] Ss-hydroxibutirat och acetoacetat är metaboliter som används som underlag i hjärtmuskler och njurbark. I hjärnan blir ketonkroppar viktiga energikällor när kroppen har uttömt sin glukosreservat.
[TOC]
Generella egenskaper
Ketogenes anses vara en mycket viktig fysiologisk eller metabolisk väg. Generellt utförs denna mekanism i levern, även om det har visats att den kan utföras i andra vävnader som kan metabolisera fettsyror.
Bildningen av ketonkroppar är den huvudsakliga metaboliska härledningen av acetyl-CoA. Denna metabolit erhålls från den metaboliska vägen som kallas ß-oxidation, som är nedbrytningen av fettsyror.
Tillgängligheten av glukos i vävnaderna där ß-oxidation inträffar bestämmer den metaboliska destinationen för acetyl-CoA. Särskilt situationer är oxiderade fettsyror nästan helt riktade mot syntesen av ketonkroppar.
Ketonkropparnas typer och egenskaper
Den huvudsakliga ketoniska kroppen är acetoacetat eller acetoättiksyra, som mestadels syntetiseras i leverceller. Från acetoacetat härstammar de andra molekylerna som utgör ketonkropparna.
Minskningen av acetoättiksyra ger upphov till D-p-hydroxibutirat, den andra ketoniska kroppen. Aceton är en svår förening att brytas ned och produceras genom en spontan dekarboxyleringsreaktion av acetoacetat (så det kräver inte ingripande av något enzym), när det finns i höga koncentrationer i blod.
Valören av ketonkroppar har arrangerats genom konvention, eftersom strikt talat ß-hydroxibutirat har inte en ketonfunktion. Dessa tre molekyler är lösliga i vatten, vilket underlättar deras blodtransport. Dess huvudfunktion är att tillhandahålla energi till vissa vävnader som skelett- och hjärtmuskler.
De enzymer som är involverade i bildandet av ketonkroppar finns främst i levern och njurarna, vilket förklarar att dessa två platser är de viktigaste producenterna av dessa metaboliter. Dess syntes sker endast och uteslutande i den mitokondriella matrisen av celler.
Det kan tjäna dig: Sporulation: I växter, i svampar och i bakterierNär dessa molekyler har syntetiserats går de till blodomloppet och adresserar vävnaderna som kräver dem, där de bryts ned tills acetyl-CoA.
Syntes av ketonkroppar
Villkor för ketogenes
Den metaboliska destinationen för acetyl-CoA från ß-oxidation beror på organismens metaboliska krav. Detta oxideras till CO2 och h2Eller genom citronsyracykeln eller syntesen av fettsyror, om metabolismen av lipider och kolhydrater är stabil i kroppen.
När kroppen behöver kolhydrater används oxalacetat för glukostillverkning (glukoneogenes) istället för att starta citronsyracykeln. Detta händer, som nämnts, när kroppen har viss oförmåga att få glukos, i fall som långvarig fasta eller närvaro av diabetes.
På grund av detta används acetyl-CoA till följd av oxidation av fettsyror för produktion av ketonkroppar.
Mekanism
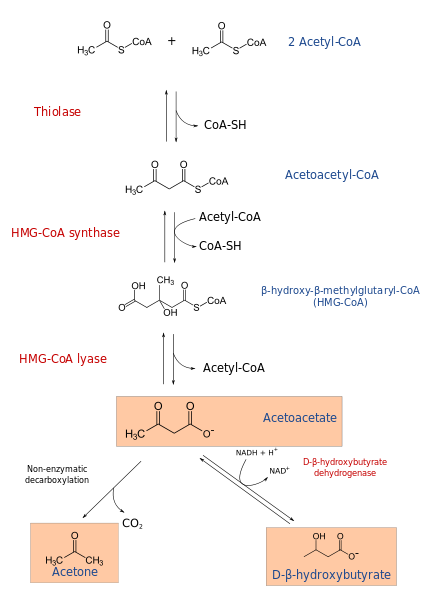
Ketogenesprocessen börjar från produkterna från ß-oxidation: acetacetyl-CoA eller acetyl-CoA. När substratet är acetyl-CoA består det första steget av kondensation av två molekyler, acetyl-CoA-transferasreaktion, för att producera acetacetyl-CoA.
The acetacetyl-COA is condensed with a third acetyl-CoA by the action of the HMG-COA synthase, to produce HMG-COA (β-hydroxy- β-methylglutaril-coa). HMG-CoA försämras till acetoacetat och acetyl-CoA genom HMG-CoA Liasa-åtgärd. På detta sätt erhålls den första ketoniska kroppen.
Acetoacetat reduceras till p-hydroxibutirat genom interventionen av p-hydroxibutiratdehydrogenas. Denna reaktion beror på NADH.
Den huvudsakliga ketoniska acetoacetatkroppen är en p-cetoacid, som upplever icke-enzymatisk dekarboxylering. Denna process är enkel och producerar aceton och co2.
Denna serie reaktioner resulterar således i ketonkroppar. Dessa är lösliga i vatten kan transporteras på ett enkelt sätt genom blodcirkulation, utan att behöva förankra till en albuminstruktur, som är fallet med fettsyror som är olösliga i vattenhaltigt medium.
Ss-oxidation och ketogenes är relaterade
Fettsyrametabolism producerar substrat för ketogenes, så dessa två sätt är funktionellt relaterade.
Acetoacetyl-CoA är en hämmare av metabolism av fettsyror, eftersom det stoppar aktiviteten hos acyl-CoA-dehydrogenas som är det första enzymet av p-oxidation. Dessutom utövar den också hämning på acetyl-CoA-överföring och HMG-CoA-syntas.
HMG-CoA-syntasenzymet, med förbehåll för CPT-I (enzym som är involverat i produktionen av acylkarnitin i p-oxidation), representerar en viktig reglerande roll i bildningen av fettsyror.
Det kan tjäna dig: Flora och fauna från Zacatecas: mer representativa arterReglering av ß-oxidation och dess effekt på ketogenes
Organismmatning reglerar en komplex uppsättning hormonsignaler. Kolhydrater, aminosyror och lipider som konsumeras i kosten deponeras i form av triacylglyceroler i fettvävnaden. Insulin, ett anaboliskt hormon, ingriper i syntesen av lipider och bildning av triacylglyceroler.
På mitokondriell nivå styrs ß-oxidation av inträde och deltagande av vissa underlag i mitokondrierna. Enzymet CPT I syntetiserar karnitin acyl från acil cytosolisk acil.
När kroppen matas ökar acetyl-CoA-karboxylas och citrat nivåerna av CPT I, medan du minskar dess fosforylering (cyklisk AMP-beroende reaktion).
Detta orsakar en ansamling av Malonil COA, som stimulerar syntesen av fettsyror och blockerar dess oxidation, vilket förhindrar att en meningslös cykel genereras.
När det gäller fasta är aktiviteten hos karboxylas mycket låg eftersom nivåerna av enzymet CPT som jag har reducerats och har också fosforylerat, aktiverat och främjar oxidationen av lipider, vilket därefter tillåter bildandet av ketonkropparna genom acetyl -COA.
Degradering
Ketonkropparna sprids utanför cellerna där de syntetiserades och transporterades till perifera vävnader av blodomloppet. I dessa vävnader kan de oxideras genom cykeln med trikarboxylsyror.
I perifera vävnader oxideras ß-hydroxibutirat till acetoacetat. Därefter aktiveras det nuvarande acetoacetatet av verkan av 3-ZOA-transfasenzymet.
Succinil-CoA fungerar som en COA-givare som blir succinat. Aktiveringen av acetoacetat sker för att förhindra succinyl-CoA.
Den resulterande aceoacetyl-CoA lider av ett tiolitiskt brott som producerar två acetyl-CoA-molekyler som är integrerade i cykeln med trikarboxylsyror, bättre känd som Krebs-cykel.
Leverceller saknar 3-kotoacil-CoA-överföringen, vilket förhindrar att denna metabolit aktiveras i dessa celler. På detta sätt är det garanterat att ketonkropparna inte oxiderar i cellerna där de producerades, utan att de kan överföras till vävnaderna där deras aktivitet krävs.
Medicinsk relevans av ketonkroppar
I människokroppen kan höga koncentrationer av ketonkroppar i blodet orsaka speciella förhållanden som kallas acidos och ketonemi.
Kan tjäna dig: sfingomyeline: vad är, struktur, funktioner, syntesTillverkningen av dessa metaboliter motsvarar katabolismen hos fettsyror och kolhydrater. En av de vanligaste orsakerna till ett tillstånd av patologisk ketogenes är den höga koncentrationen av ättiksdikarbonerade fragment som inte bryts ned på oxidationsvägen för tricarboxylsyror.
Som en följd av detta finns det en ökning av nivåerna av blod ketonkroppar över 2 till 4 mg/100 N och deras närvaro i urin. Detta innebär störningen av mellanhandsmetabolismen hos dessa metaboliter.
Vissa defekter i hypofysen neuroglandulära faktorer som reglerar nedbrytning och syntes av ketonkropparna, tillsammans med störningar i kolväte metabolism, är orsaken till tillståndet för hypercetonemi.
Diabetes mellitus och ackumulering av ketoniska kroppar
Diabetes mellitus (typ 1) är en endokrin sjukdom som orsakar en ökning av produktionen av ketonkroppar. Insulin Otillräcklig produktion inaktiverar glukos till muskler, lever och fettvävnad, vilket samlas i blod.
Celler i frånvaro av glukos börjar processen med glukoneogenes och nedbrytning av fett och proteiner för att återställa deras ämnesomsättning. Som en följd minskar och ökar oxalacetatkoncentrationerna oxidation av lipider.
En acetyl-CoA-ackumulering inträffar, som i frånvaro av oxalacetat inte kan följa citronsyrans väg och sedan orsaka de höga produktioner av ketonkropparna, karakteristiska för denna sjukdom.
Ackumuleringen av aceton upptäcks av dess närvaro i urinen och i andetaget av människor som presenterar detta tillstånd och är i själva verket ett av symtomen som indikerar manifestationen av denna sjukdom.
Referenser
- Blázquez Ortiz, c. (2004). Ketogenes i astrocyter: karakterisering, reglering och möjlig cytoprotektivt papper (Dissertation Doctoral, Complutense University of Madrid, Publications Service).
- Devlin, t. M. (1992). Lärobok för biokemi: Med kliniska korrelationer.
- Garrett, R. H., & Grisham, c. M. (2008). Biokemi. Thomson Brooks/Cole.
- McGry, J. D., Mannaerter, g. P., & Foster, D. W. (1977). En möjlig roll för malonyl-CoA i regleringen av leverfettsyraoxidation och ketogenes. Journal of Clinical Investigation, 60(1), 265-270.
- Melo, v., Ruiz, V. M., & Cuamatzi, eller. (2007). Biokemi för metaboliska processer. Reverte.
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger principer för biokemi. Macmillan.
- Pertierra, a. G., Gutiérrez, c. V., & Andra, c. M. (2000). Grundläggande metaboliska biokemi. Redaktion.
- Voet, D., & Voet, J. G. (2006). Biokemi. Ed. Pan -amerikansk medicin.
- « Kontinental frukost vad är och vilka livsmedel den inkluderar?
- Kalciumperoxid (CAO2) egenskaper, risker och användningar »