

Fettsyrorstruktur, typer, funktioner, biosyntes
- 2208
- 509
- PhD. Emil Svensson
De fettsyror De är organiska makromolekyler härrörande från kolväten, som består av långa kedjor av kol- och väteatomer som är hydrofoba (är feta -lösliga) och är den strukturella grunden för fetter och lipider.
De är mycket olika molekyler som skiljer sig från varandra med längden på deras kolvätekedjor och närvaro, antal, position och/eller konfiguration av deras dubbelbindningar.
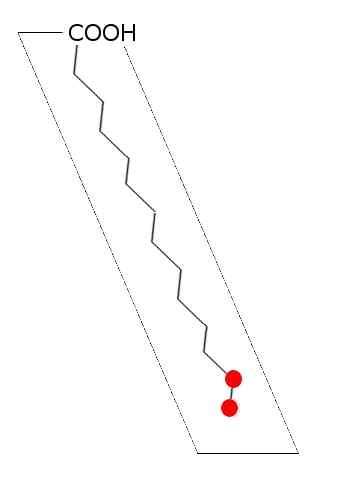
 Allmänt schema för mättad fettsyra (källa: Laghi.L [CC BY-SA 3.0 (http: // Creativecommons.Org/licenser/BY-SA/3.0/)] via Wikimedia Commons)
Allmänt schema för mättad fettsyra (källa: Laghi.L [CC BY-SA 3.0 (http: // Creativecommons.Org/licenser/BY-SA/3.0/)] via Wikimedia Commons) I lipider av djur, växter, svampar och mikroorganismer som bakterier och jästar har mer än 100 slags olika fettsyror beskrivits och de anses vara specifika arter och vävnad i de flesta levande varelser.
De oljor och fetter som människan konsumerar dagligen är av djur eller växt ursprung, består mestadels av fettsyror.
[TOC]
Introduktion
 Smör, bland annat, består i huvudsak av fettsyror (källa: Africa Studio, via Pixabay.com)
Smör, bland annat, består i huvudsak av fettsyror (källa: Africa Studio, via Pixabay.com) Fettsyramolekyler utför viktiga funktioner på cellnivå, vilket gör dem till viktiga komponenter och eftersom vissa av dem inte kan syntetiseras av djur måste de få dem från kosten.
Fettsyror är sällsynta som fria arter i cellcytosol, så de är i allmänhet en del av andra molekylära konjugat såsom:
- Lipider, i biologiska membran.
- Triglycerider eller fettsyraestrar, som fungerar som reserv i växter och djur.
- Vax, som är solida estrar av fettsyror och alkoholer med lång kedja.
- Andra liknande ämnen.
Hos djur lagras fettsyror i cytoplasma i celler som små droppar fett som består av ett komplex som kallas triacylglycerol, som inte är något annat än en glycerolmolekyl som den har anslutit sig till, i var och en av dess kol, en fettsyrakedja av Ester Type Binds.
Medan bakterier har korta och vanligtvis enomättade fettsyror, är det i naturen vanligt.
Strukturera
Fettsyror är amfipatiska molekyler, det vill säga de har två kemiskt definierade regioner: en hydrofil polär region och en hydrofob apolär region.
Den hydrofoba regionen består av en lång kolvätekedja som i kemiska termer inte är särskilt reaktiv. Den hydrofila regionen består å andra sidan av en karboxylgrupp (-COH) -terminal, som uppför sig som en syra.
Denna terminala karboxylgrupp eller karboxylsyra är joniserad i lösning, den är mycket reaktiv (kemiskt sett) och är mycket hydrofil, så den representerar ett kovalent korsningsställe mellan fettsyra och andra molekyler.
Längden på kolvätekedjorna av fettsyror har vanligtvis till och med antal kolatomer, och detta är nära besläktat med den biosyntetiska processen genom vilken de produceras, eftersom deras tillväxt inträffar från kolparpar.
De vanligaste fettsyrorna har kedjor mellan 16 och 18 kolatomer och djur, dessa kedjor är inte grenade.
Klassificering
Fettsyror klassificeras i två stora grupper beroende på arten av länkarna som komponerar dem, det vill säga efter förekomsten av enkla bindningar eller dubbelbindningar mellan kolatomerna i deras kolvätekedjor.
Således finns det mättade och omättade fettsyror.
- Mättade fettsyror har bara enkla kol - kolbindningar och alla deras kolatomer är "mättade" eller kopplade till vätemolekyler.
- Omättade fettsyror har en eller flera dubbla kol - kolbindningar och inte alla dessa är fästa vid en väteatom.
Omättade fettsyror är också uppdelade enligt antalet omättnad (dubbelbindningar) i enomättade, de med en enda dubbelbindning och fleromättade, som har mer än en.
Mättade fettsyror
De har vanligtvis mellan 4 och 26 kolatomer förenade med enkla länkar. Dess smältpunkt är direkt proportionell mot kedjans längd, det vill säga till dess molekylvikt.
Fettsyrorna som har mellan 4 och 8 kol är flytande vid 25 ° C och är de som utgör ätliga oljor, medan de med mer än 10 kolatomer är solida.
Bland de vanligaste är laurinsyran, som är riklig i palm- och kokosnötsfrön; Palmitinsyra, som finns i handflatan, kakao och grissmör, och stearinsyra, som finns i kakao och hydrerade oljor.
De är fettsyror med mycket mer stabilitet än omättade fettsyror, särskilt inför oxidation, åtminstone under fysiologiska förhållanden.
Tack vare det faktum att enkla kolbindningar kan rotera gratis.
Omättade fettsyror
Dessa fettsyror är mycket reaktiva och benägna till mättnad och oxidation. De är vanliga i växter och marina organismer. De som har en enda dubbelbindning kallas enomättad eller monoenoik, medan de som har mer än två är kända som polyenik eller fleromättade.
Kan tjäna dig: DNA -polymerasNärvaron av dubbelbindningar är vanligt bland kolatomer mellan position 9 och 10, men detta betyder inte att monouinsättade fettsyror inte finns med en omättning i en annan position.
Till skillnad från mättade, omättade fettsyror listas inte från den terminala karboxylgruppen, utan enligt positionen för den första dubbelbindningen c - c. Således är dessa uppdelade i två grupper, omega-6 eller ω6 och omega-3 eller ω3.
Omega-6-syror har den första dubbelbindningen i kol nummer 6 och omega-3-syror har det i kol nummer 3. N valöret ω ges av dubbelbindningen närmast den slutliga metylgruppen.
Dubbel länkar finns också i två geometriska konfigurationer kända som "Cis " och "trans".
De flesta naturliga omättade fettsyror har konfiguration "Cis”Och dubbelbindningarna av fettsyror som finns i kommersiella fetter (hydrerade) finns i "trans".
I fleromättade fettsyror separeras vanligtvis två dubbelbindningar från varandra åtminstone av en metylgrupp, det vill säga en kolatom fäst vid två väteatomer.
Funktioner
Fettsyror har flera funktioner i levande organismer och, som nämnts ovan, en av deras väsentliga funktioner som en väsentlig del av lipider, som är de viktigaste komponenterna i biologiska membran och en av de tre vanligaste biomolekylerna i organismer i organismer som lever i samband med proteiner och kolhydrater.
De är också utmärkta energisubstrat tack vare vilka stora mängder energi erhålls i form av ATP och andra mellanhandsmetaboliter.
Med tanke på det faktum att djur, till exempel, inte kan lagra kolhydrater, representerar fettsyror den huvudsakliga källan för energilagring som kommer från oxidation av överskott av sockerarter.
Kort -kedjor mättade fettsyror i kolon deltar i stimulering av vattenabsorption och natrium-, klorid- och bikarbonatjoner; Dessutom har de funktioner i produktionen av slem, i spridningen av kolonocyter (kolonceller), etc.
Omättade fettsyror är särskilt rikliga i ätliga oljor av växtens ursprung, som är viktiga i kosten för alla människor.
 Oljorna vi konsumerar dagligen är fettsyror (källa: Steve.com)
Oljorna vi konsumerar dagligen är fettsyror (källa: Steve.com) Andra deltar som ligander av vissa proteiner med enzymatiska aktiviteter, så de är viktiga med avseende på deras effekter på energimetabolismen i cellerna där de finns.
Biosyntes
Nedbrytningen av fettsyror kallas p-oxidation och förekommer i mitokondrierna i eukaryota celler. Biosyntes, tvärtom, förekommer i cytosolen hos djurceller och i kloroplaster (fotosyntetiska organeller) av växtceller.
Det är en acetyl-CoA, malonyl-CoA och NADPH-process, den förekommer i alla levande organismer och hos "överlägsna" djur som däggdjur. Till exempel är det mycket viktigt i lever- och fettvävnader, liksom i bröstkörtlar.
NADPH som används för denna rutt är huvudsakligen en produkt av de NADP-beroende oxidationsreaktionerna från pentosfosfatvägen, medan acetyl-CoA kan komma från olika källor, till exempel från den oxidativa decarboxylering av pyruvatet, av Krebs och p- oxidation av fettsyror.
Biosyntesvägen, som ß-oxidation, är starkt reglerad i alla celler av alestheriska effektorer och kovalenta modifieringar av enzymer som deltar i förordningen.
-Malonyl-CoA-syntes
Rutten börjar med bildandet av en metabolisk mellanhand känd som malonyl-CoA från en acetyl-CoA-molekyl och katalyseras av ett multifunktionellt enzym som kallas acetyl-CoA-karboxilasa.
Denna reaktion är en reaktion på tillägg till en karboxylmolekyl (-COH, karboxylering) beroende på biotin och förekommer i två steg:
- Först överföringen, ATP-beroende, av en karboxyl härrörande från bikarbonat (HCO3-) till en biotinmolekyl som finns som en protes (icke-protein) associerad med acetyl-CoA-karboxylas.
- Därefter överförs CO2 till acetyl-CoA och malonyl-CoA produceras.
-Ruttreaktioner
Hos djur inträffar bildningen av kolhydratkedjor av fettsyror ytterligare genom sekventiella kondensationsreaktioner katalyserade av ett multikrititiskt och multifunktionellt enzym som kallas fettsyrasyntas.
Detta enzym katalyserar kondensationen av en acetyl-CoA-enhet och multonil-CoA-molekyler som produceras från reaktionen av karboxylasacetyl-CoA, process under vilken en CO2-molekyl frigörs för varje malonyl-CoA som den lägger till.
Växande fettsyror sterifieras till ett protein som kallas "acylbärarprotein" eller ACP, som bildar tioéer med acylgrupper. I OCH. coli Detta protein är en 10 kDa polypeptid, men hos djur är det en del av fettsyrakomplexsyntas.
Kan tjäna dig: noxaBrottet av dessa bindningar tioéster släpper ut stora mängder energi, vilket gör det möjligt, termodynamiskt sett, förekomsten av kondensationssteg på den biosintetiska vägen.
Fettsyrakomplex syntas
I bakterier motsvarar syntas-syraaktiviteten faktiskt sex oberoende enzymer som använder acetyl-CoA och malonyl-CoA för att bilda fettsyror och med vilka sex olika enzymatiska aktiviteter är associerade.
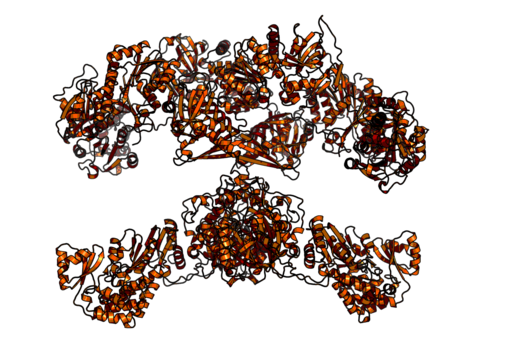 Homodimeriska och multifunktionella komplexa fettsyrasyntas av djur (källa: Boehringer Ingelheim [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons)
Homodimeriska och multifunktionella komplexa fettsyrasyntas av djur (källa: Boehringer Ingelheim [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons) Hos däggdjur, å andra sidan, är fettsyrasyntas ett multifunktionellt homodimiskt enzymatiskt komplex på cirka 500 kDa molekylvikt, som har sex olika katalytiska aktiviteter och med vilket syrabärarproteinet är associerat.
Steg 1: Prowter -reaktion
Tiolgrupper i cysteinavfall som ansvarar för föreningen av metaboliska mellanhänder till AVS -enzymet måste laddas, före syntesens början, med nödvändiga syrogrupper.
För att göra detta överförs acetyl-CoA-acetylgruppen till Tiol (-SH) -gruppen i en av cysteinresterna i ACP-underenheten för fettsyrasyntas. Denna reaktion katalyseras av ACP-akil-transferasunderenheten.
Acetylgruppen överförs sedan från ACP till en annan cysteinrest i det katalytiska stället för en annan enzymatisk underenhet i komplexet som kallas ß-cetoacyl-ACP-SINTA. Således är det enzymatiska komplexet "korn" för att börja syntes.
Steg 2: Överföring av malonyl-CoA-enheter
Malonyl-CoA som produceras av acetyl-CoA-karboxilasa överförs till tiolgruppen i ACP och under denna reaktion går COA-delen förlorad. Reaktionen katalyseras av Malonil-ACP-transferasunderenheten för fettsyrasyntaskomplexet, som sedan producerar Malonil-ACP.
Under denna process är den maloniska gruppen kopplad till ACP och ß-Cotoacil-ACP-SITA genom en esterlänk respektive en annan sulfidl.
Steg 3: Kondensation
Den ß-zoacil-ACP-suntasenzymet katalyserar överföringen av acetylgruppen som gick med honom i passagen av "korn" mot kol 2 i Malonilo-gruppen som i föregående steg överfördes till ACP.
Under denna reaktion frisätts en CO2-molekyl som kommer från malonylen, vilket motsvarar CO2 som bikarbonatet har bidragit med i karboxyleringsreaktionen hos karboxylasacetyl-CoA. Sedan inträffar acetoacetyl-ACP.
Steg 4: Minskning
P-Zoacil-ACP-reduktas-underenheten katalyserar reduktionen av NADPH-beroende ACEOACETYL-ACP.
Steg 5: Dehydrering
Detta steg bildar trans-a, ß-acyl-ACP eller ∆2-inoRAGADO-ACIL-ACP (crratonil-ACP), produkt från dehydrering av D-p-hydroxybutiril-ACP genom handling av den arga underenheten ACP -Hydratasa.
Senare reduceras cratonil-ACP till butiril-ACP av en reaktion beroende på NADPH katalyserad av den arga-AC-reduktas subenheten. Med denna reaktion är den första av sju cykler som behövs för att producera palmitail-ACP, som är föregångaren till nästan alla fettsyror är klar.
Hur följer efterföljande kondensationsreaktioner?
Butirilo-gruppen överförs från ACP till tiolgruppen av en cysteinrest i ß-cethoacil-ACP-SITA, så att ACP kan acceptera en annan malonisk grupp från Malonyl-CoA.
På detta sätt är reaktionen som händer kondensationen av Malonil-ACP med buturil-p-kotoacil-ACP-SINTA, vilket ger upphov till ß-zotohexanoil-ACP + CO2.
Palmitail-ACP som uppstår från de efterföljande stegen (efter tillägget av ytterligare 5 maloniska enheter) kan frisättas som fri palmitinsyra tack vare aktiviteten hos enzymtioesteraset kan det överföras till COA eller införliva i fosfatidsyran för för Syntesvägen för fosfolipider och triacylglycerider.
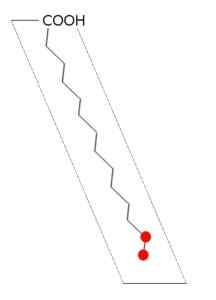
 Palmitinsyrastruktur (källa: Andel, via Wikimedia Commons)
Palmitinsyrastruktur (källa: Andel, via Wikimedia Commons) Fettsyrasyntaset av de flesta organismer stannar i syntesen av palmitail-ACP, med tanke på att det katalytiska stället för p-cetoacyl-ACP-syntasunderenheten har en konfiguration som endast fettsyror av den längden kan rymmas.
Hur är fettsyror med udda antal kolatomer?
Dessa är relativt vanliga i marina organismer och syntetiseras också av ett komplex fettsyrasyntas. Men "korn" -reaktionen inträffar med en längre molekyl, propionyl-ACP, av tre kolatomer.
Var och hur är fettsyrorna i längre kedjor?
Palmitinsyra, som kommenterat, fungerar som en föregångare för många mättade och omättade fettsyror av längre kedjor. "Förlängning" -processen för fettsyror sker i mitokondrier, medan införandet av omättning sker väsentligen i endoplasmatisk retikulum.
Många organismer förvandlar sina mättade fettsyror till omättade som en anpassning till låga miljötemperaturer, eftersom detta gör att de kan hålla fusionspunkten för lipider under omgivningstemperaturen.
Fettsyraegenskaper
Många av egenskaperna hos fettsyror beror på deras kedjelängd och närvaron och antalet omättnad:
Kan tjäna dig: Abiotisk synteseori: Huvudegenskaper- Omättade fettsyror har lägre smältpunkter än mättade fettsyror av samma längd.
- Längden på fettsyror (antalet kolatomer) är omvänt proportionell mot molekylens fluiditet eller flexibilitet, det vill säga de mest "korta" molekylerna är mer flytande och vice versa.
I allmänhet består flytande fettämnen av fettsyror med korta kedjor och med omättnad närvaro närvaro.
Växter har riklig mängd omättade fettsyror, liksom djur som lever vid mycket låga temperaturer, eftersom dessa, som komponenter i lipider som finns i cellmembran, ger dem större flytande under dessa förhållanden.
Under fysiologiska förhållanden.
Närvaron av dubbelbindningar i fettsyror förknippade med lipidmolekyler har direkta effekter på graden av "förpackning" som de kan ha i de membran som de tillhör och med det har de också effekter på membranproteiner.
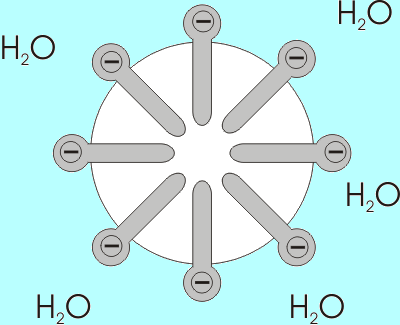 Exempel på bildandet av en myla av fettsyror med karboxylgrupper exponerade mot den vattenhaltiga miljön (källa: Benutzer: Anderl [CC BY-SA 3.0 (http: // Creativecommons.Org/licenser/BY-SA/3.0/)] via Wikimedia Commons)
Exempel på bildandet av en myla av fettsyror med karboxylgrupper exponerade mot den vattenhaltiga miljön (källa: Benutzer: Anderl [CC BY-SA 3.0 (http: // Creativecommons.Org/licenser/BY-SA/3.0/)] via Wikimedia Commons) Lösligheten hos fettsyror minskar när längden på kedjan ökar, så de är omvänt proportionella. I vattenhaltiga och lipidblandningar är fettsyror associerade i strukturer som kallas miceller.
En micella är en struktur där de alifatiska kedjorna av fettsyror är "inneslutna", och därmed "utvisande" alla vattenmolekyler och på vars yta finns karboxylgrupper.
Nomenklatur
Nomenklaturen för fettsyror kan vara något komplex, särskilt om man hänvisar till de vanliga namnen de får, som ofta är relaterade till viss fysikalisk -kemisk egendom, med platsen där de är eller andra egenskaper.
Många författare anser att som tack vare den terminala karboxylgruppen dessa molekyler är joniserade till fysiologiskt pH, bör man hänvisa till dem som "karboxilat" med avslutningen "Ato ".
Enligt IUPAC -systemet. Kedjeterterminalmetylen innehåller kolatom Ω.
I allmänhet får de i den systematiska nomenklaturen namnet på "föräldrakolvätet (kolväte med samma antal kolatomer) och deras avslutning ersätts"antingen" förbi "Oico", Om det är en omättad fettsyra läggs avslutningen "Dimma".
Tänk till exempel på fallet med en fettsyra med 18 kolatomer (C18):
- Eftersom kolväte med samma mängd kolatomer kallas oktadekano kallas mättad syra "Oktadekansyra" Nåväl "Oktadekano”Och dess vanliga namn är stearinsyra.
- Om du har en dubbelbindning mellan några par kolatomer i dess struktur, är det känt som "Oktadekensyra"
- Om du har två Double C Res C - C, kallas det "Octadecadienoic Acid" Och om du har tre "Oktadekatriensyra".
Om du vill sammanfatta nomenklaturen, använd 18: 0 för 18 -kolfettsyra och ingen dubbelbindning (mättad) och beroende på graden av omättnad, istället för noll skrivs 18: 1 för en molekyl med en molekyl med en omättnad, 18: 2 för en med två omättnad och så vidare.
Om du vill ange mellan vilka kolatomer som är dubbelbindningar i omättade fettsyror, används ∆ -symbolen med en numerisk undersökning som indikerar platsen för omättnad och prefix "Cis"antingen "trans", beroende på konfigurationen av detta.
Referenser
- Badui, s. (2006). Matkemi. (OCH. Quintanar, Ed.) (4: e upplagan.). Mexiko D.F.: Pearson Education.
- Garrett, R., & Grisham, c. (2010). Biokemi (4: e upplagan.). Boston, USA: Brooks/Cole. Cengage Learning.
- Mathews, C., Van Holde, K., & Ahern, K. (2000). Biokemi (3: e upplagan.). San Francisco, Kalifornien: Pearson.
- Murray, r., Bender, D., Botham, K., Kennelly, s., Rodwell, V., & Weil, s. (2009). Harpers illustrerade biokemi (28: e upplagan.). McGraw-Hill Medical.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger principer för biokemi. Omega -utgåvor (5: e upplagan.).
- Rawn, j. D. (1998). Biokemi. Burlington, Massachusetts: Neil Patterson Publishers.
- Tvrzicka, E., Kremmyda, L., Stankova, b., & Zak, till. (2011). Fettsyror som biokompositioner: deras roll i mänsklig metabolism, hälsa och sjukdom- en översyn. Del 1: Klassificering, dietkällor och biologiska funktioner. Biomed pap med fac univ palacky olomouc tjeckiska repub, 155(2), 117-130.
- « Glykoproteins struktur, funktioner, klasser och exempel
- Effekterna av att tillhöra digitalt medborgarskap »