

Cytosinstruktur, funktioner, egenskaper, syntes
- 1101
- 215
- Erik Eriksson
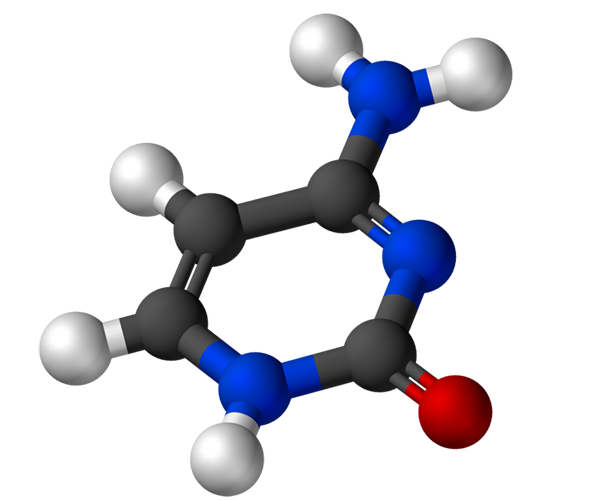
De Cytosin Det är en pyrimidin kvävebas, som tjänar för biosyntes av cytidin-5'-monofosfat och dexicitidin-5'-monofosfat. Dessa föreningar tjänar för biosyntes, respektive från deoxyribonukleinsyra (DNA) och ribonukleinsyra (RNA). DNA lagrar genetisk information och RNA har olika funktioner.
I levande varelser finns inte cytosin fritt, men vanligtvis ribonukleotider eller deoxyribonukleotider. Båda typerna av förening har en fosfatgrupp, en ribos och en kvävebas.
Källa: VESPROM [Public Domain] Kol 2 i ribosen har en oxhydrillo-grupp (-OH) i ribonukleotiderna och en väteatom (-H) i deoxyribonukleotiderna. Beroende på antalet närvarande fosfatgrupper finns det citidín-5'-monofosfat (CMP), citidín-5'-difosfat (CDP) och citidín-5'-triffosfat (CTP) (CTP).
Desoxygenerade ekvivalenter kallas desoxicitidin-5'-monofosfat (DCMP), desoxicitidin-5'-difosfat (DCDP) och desoxicitidin-5'-triffosfat (DCTP).
Cytosin deltar i sina olika former, i olika funktioner, såsom biosyntes av DNA och RNA, biosyntesen av glykoproteiner och regleringen av uttrycket av genen.
[TOC]
Struktur och egenskaper
Cytosinet, 4-amino-2-hydroxipirimidin, har den empiriska formeln C4H5N3Eller vars molekylvikt är 111,10 g/mol och renas som ett vitt pulver.
Strukturen för cytosin är en heterocyklisk, aromatisk och plan ring. Den maximala absorbansvåglängden (ʎMax) Det är 260 nm. Fusionstemperaturen för cytosinet överstiger 300 ºC.
För att bilda en nukleotid binder cytosin kovalent, genom kväve 1, med hjälp av medel. Carbon 5 'förstärks med en fosfatgrupp.
Biosyntes
Pyrimidin nukleotidbiosyntes har en vanlig väg, som består av sex steg katalyserade av enzymer. Vägen börjar med karbamoil-fosfatbiosyntes. I prokaryoter finns det bara ett enzym: karbamoilfosfatsyntas. Detta ansvarar för syntesen av pyrimidiner och glutamin. I eukaryoter finns det karbamoilfosfatsyntas I och II, som är ansvariga för glutamin- och pyrimidinbiosyntes.
Det andra steget består av bildandet av N-Carbamoilaspartato, från kolfosfat och aspartat, som katalyseras av transpartat Transcabamoilasa (Atcasa).
Det tredje steget är syntesen av L-dihydrotoat, som ger stängningen av pyrimidinringen. Detta steg katalyseras av dihydrootas.
Det fjärde steget är bildandet av orotato, som är en redoxreaktion katalyserad av dihydroorotatdehydrogenas.
Det femte steget består av bildandet av goldylat (OMP) med användning av fosforibosylpyrofosfat (PRPP) som ett substrat och fosforibosyl -orotattransferaset som katalysator som katalysator.
Det sjätte steget är bildandet av uridilaato (Uridin-5'-monofosfat, UMP), reaktion katalyserad av ett OMP-decarboxylas.
Det kan tjäna dig: flora och fauna från HidalgoFöljande steg består av fosforylering av UMP, katalyserade av kinaser, för att bilda UTP, och överföring av en aminogrupp från glutamin till UTP för att bilda CTP, reaktion katalyserad av CTP -syntetas.
Biosyntesreglering
Hos däggdjur sker reglering vid karbamoilfosfatsyntas.
CARBAMOIL SYNTASA II regleras av negativ feedback. Dess regulatorer, UTP och PRPP, är respektive hämmare och aktivator för detta enzym.
I vävnader som inte är lever är karbamoilfosfatsyntas II den enda källan till karbamoilfosfat. Medan i levern, under förhållanden med överskott av ammoniak, producerar karbamoilfosfatsyntas I, i mitokondrierna, karbamoilfosfat, som transporteras till cytosolen, varifrån den kommer in i biosyntesvägen av pyrimidiner.
En annan regleringspunkt är OMP-diskarboxylas, som regleras av konkurrenskraftig hämning. Produkten från dess reaktion, Ump, tävlar med OMP för bindningsstället i OMP-descarboxilasa.
Pyrimidiner, som cytosin, återvinns
Återvinning av pyrimidiner har som en funktion återanvändning av pyrimidiner utan behov av novo -biosyntes och undvik den nedbrytande vägen. Återvinningsreaktionen katalyseras av fosforibosyltransferaspyrimimidin. Den allmänna reaktionen är som följer:
Pyrimidin + PrPP -> nukleosidpyrimidin 5 '-Monofosfat + PPI
I ryggradsdjur finns fosforibosyltransferaspyrimimidin i erytrocyter. Pyrimidinsunderlaget för detta enzym är uracil, timina och orotato. Cytosin återvinns indirekt från uridin-5'-monofosfat.
Funktion i DNA -biosyntes
Under DNA -replikering kopieras informationen i DNA i DNA genom ett DNA -polymeras.
RNA -biosyntes behöver deoxynukleotider tryfosfat (DNTP), nämligen: dexitimid tryfosfat (DTTP), dexicitidy triffosfat (DCTP), de -alxyxyadenin tryphyse (DATP) och dexiguanin tryfosfat (DGTP), DGTP). Reaktionen är:
(DNA)n avfall + Dntp -> (DNA)N+1 avfall + ppi
Oorganisk pyrofosfat (PPI) hydrolys ger energi för biosyntes av RNA.
Funktion i stabiliseringen av DNA: s struktur
I den dubbla dnus -spiralen är en purin, av en kedja, kopplad till pyrimidin, i motsatt kedja, av vätebindningar. Således är cytosin alltid kopplad till guanin av tre vätebindningar: adenin är kopplad till timin av två vätebindningar.
Det kan tjäna dig: Biologisk betydelseVätebindningar bryts när en renad nativ DNA -lösning vid pH 7 utsätts för temperaturer större än 80 ºC. Detta får den dubbla DNA -propellen att bilda två separata kedjor. Denna process kallas denaturering.
Temperaturen vid vilken 50% av DNA denatureras kallas smälttemperaturen (TM). DNA -molekyler vars guanin och cytosin.
Ovanstående är det experimentella testet som ett större antal vätebindningar bättre stabiliserar de nativa DNA -molekylerna.
Funktion av rika regioner i cytosin i DNA
Nyligen konstaterades att DNA från den mänskliga cellkärnan kan anta interkalerade motivstrukturer (IM). Dessa strukturer produceras i cytosinregioner.
IM -strukturen består av fyra DNA -kedjor, till skillnad från det klassiska dubbla propeller -DNA som har två kedjor. Mer specifikt är två parallella duplexkedjor isär i en antiparallelorientering och hålls tillsammans av ett par hemiprotonerade cytosiner (C: C: C: C: C: C: C: C: C: C: C: C: C: C: C: C: C: C: C: C: C+).
I det mänskliga genomet finns IM -strukturer i regioner som promotorer och telomerer. Antalet IM -strukturer är högre under G1/S -fasen i cellcykeln, där transkriptionen är hög. Dessa regioner är proteinigenkänningsplatser involverade i aktiveringen av transkriptionella maskiner.
Å andra sidan, i regionerna som är rika på guaninbaser (c) i följd, tenderar DNA att anta propellerform A, under dehydratiseringsförhållanden. Denna form är typisk för RNA och dubbla DNA-ANN-blandade band under transkription och replikering, och vid vissa tider när DNA är kopplat till protein.
Det har visats att regionerna med på varandra följande cytosinbaser skapar en elektropositiv lapp i DNA: s huvudspalt. Därför tros det att dessa regioner binder till proteiner, som predisponerar vissa genomiska regioner av genetisk bräcklighet.
Funktion i RNA -biosyntes
Under transkript kopieras informationen i DNA i RNA genom ett PON -polymeras -RNA. RNA -biosyntes behöver tryposfat nukleosider (NTP), nämligen: tryphyse citidin (CTP), tryposfaturidin (UTP), adenin tryposfat (ATP) och trypefosfat guanin (GTP). Reaktionen är:
Kan tjäna dig: Flora och Fauna of Campeche: Representativa arter(RNA)n avfall + Ntp -> (RNA)N+1 avfall + ppi
Oorganisk pyrofosfat (PPI) hydrolys ger energi för biosyntes av RNA.
Funktion i biosyntes av glykoproteiner
Den sekventiella överföringen av hexoser för att bilda oligosackariderna, O-ligater till proteiner, händer från nukleotidprekursorer.
I ryggraden består det sista steget i biosyntesen av O-ligerade oligosackarider i tillsats av två sinalsyraavfall (N-acetylneuramin) från en föregångare av citidin-5'-monofosfat (CMP). Denna reaktion produceras i transsäcken till Golgi.
Cytosin- och kemoterapibehandlingar mot cancer
Tetrahydrofolatsyra (FH4) är en gruppkälla3, Och det är nödvändigt för DTMP -biosyntes från dumpning. FH2 är också bildad. FH2 -reduktion till FH4 kräver ett folat- och NADPH -reduktas. Vissa folatreduktashämmare, såsom aminopterin och metretotrexat används i cancerbehandlingar.
Metotrexan är en konkurrenskraftig hämmare. Folatreduktaset går med 100 gånger mer affinitet till denna hämmare än till dess underlag. Aminapterin fungerar på liknande sätt.
Hämningen av folatreduktas hindrar indirekt DTMP -biosyntes, och därför den för DCTP. Direkt hämning sker genom hämmare av det timidilatsyntetaset, som katalyserar DTMP från dumpning. Dessa hämmare är 5-fluorouracil och 5-fluoro-2-dioxiuridin.
Till exempel är 5-fluoroacil inte en hämmare men blir först, återvinningsvägen, i deoxyurid mofosfat D (FDUMP), som binder till timidilatsyntesen och hämmar det.
Ämnen analoga med glutamin, azaserin och akivicin, hämmar amidotransferas glutamin. Azarin var ett av de första upptäckta ämnena som fungerar som självmordsinaktiverande.
Referenser
- Assi, h.TILL., Garavís, m., González, c., Och Damha, m.J. 2018. I-MOTIF DNA: Strukturella egenskaper och betydelse för cellbiologi. Nuclei Acids Research, 46: 8038-8056.
- Bohinski, r. 1991. Biokemi. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, t.M. 2000. Biokemi. Redaktionell återvändelse, Barcelona.
- Lodish, h., Berk, A., Zipurski, s.L., Matsudaria, s., Baltimore, D., Darnell, J. 2003. Cellulär och molekylärbiologi. Redaktör Medica Panamericana, Buenos Aires, Bogotá, Caracas, Madrid, Mexiko, Sāo Paulo.
- Nelson, D. L., Cox, m. M. 2008. Lehninger-principer för biokemi. W.H. Freeman, New York.
- Voet, D. och Voet, J. 2004. Biokemi. John Wiley och Sons, USA.