

Cardiolipin -struktur, syntes, funktioner
- 1720
- 508
- Lars Eriksson
De Kardiolipin, Även känd som defathidylglycerol, han är en lipid av familjen av glyceofosfolipider och gruppen av polyglyfolipider. Det finns i det mitokondriella membranet av eukaryota organismer, i plasmamembranet i många bakterier och även i vissa bågar.
Det upptäcktes av Pangborn 1942 från analysen av membranlipiderna i hjärtvävnaden i nötkreaturen. Dess struktur föreslogs 1956 och kemisk syntes ägde rum cirka tio år senare.
Cardiolipin -struktur (källa: edgar181 [public domain] via Wikimedia Commons) Vissa författare anser att deras närvaro är begränsad till ATP-producerande membran, liksom fallet med mitokondrier i eukaryoter, plasmamembran i bakterier och vätosomer (typ-mmitokondrier) i vissa protister.
Det faktum att kardiolipin befinner sig i mitokondria och i plasmamembranet på bakterier har använts för att förstärka grunden för endosimbiotic teori, som fastställer att mitokondria uppstod uppstod hos progenitorcellerna för eukaryoter genom fagocytos av ett bakterium, som då han var beroende av. cellen och vice versa.
Dess biosyntetiska väg hos djur beskrevs mellan 1970 och 1972 och därefter visades det att det är samma väg som förekommer i växter, jäst, svampar och ryggradslösa djur. Det är inte en mycket riklig lipid, men cellerna kräver att den fungerar korrekt.
Betydelsen av denna fosfolipid för mitokondrier och därför för cellmetabolism är tydlig när den defekta funktionen av metaboliska vägar associerade med den ger en mänsklig patologi som kallas Barths syndrom (kardiooskeletal myopati).
[TOC]
Strukturera
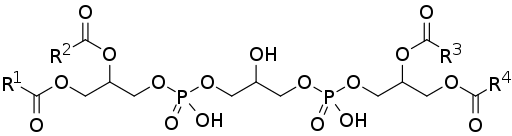
Cardiolipin eller defospathidylglycerol består av två fosfatidsyra (enklare fosfolipid) kopplade till varandra genom en glycerolmolekyl.
Fosfatidinsyra, en av de vanliga mellanhänderna på de biosyntetiska vägarna för andra fosfolipider, består av en 3-fosfatglycerolmolekyl, till vilka två kedjor av fettsyror förstörs i positionerna för kol 1 och 2, så det är också känt som 1.2 -Diacylglycerol 3-fosfat.
Kan tjäna dig: sekundär struktur av proteiner: egenskaperDärför består kardiolipin av tre glycerolmolekyler: en central glycerol, tillsammans med en fosfatgrupp i kol 1, en annan fosfatgrupp i kol 3 och en hydroxylgrupp i kol 2; och två "laterala" glyceroler.
De två "laterala" glycerolmolekylerna binder till den centrala molekylen med "Glycerolbroar" genom sina kol i position 3. I kol av positionerna 1 och 2 har två kedjor av fettsyror och variabel mättnad försttas.
Cardiolipin är en lipid som kanske eller inte kan bilda BICAPA, beroende på närvaro eller frånvaro av divalent katjoner. Detta är relaterat till det faktum att det är en symmetrisk molekyl, vilket gör det viktigt i membran som är ansvariga för energitransduktionsprocesser.
Liksom de andra lipiderna i gruppen av polyglyfosfolipider har kardiolipin flera hydroxylgrupper som kan tjäna för föreningen av fettsyror. Därför har den flera positionella stereoisomerer.
Dess fettsyror
Olika studier har fastställt att fettsyror kopplade till de laterala glycerolmolekylerna av kardiolipin vanligtvis är omättade, men graden av omättnad har inte fastställts.
Då kan sådana fettsyror ha mellan 14 och 22 kolhydrater och från 0 till 6 dubbelbindningar. Detta och det faktum att cardiolipin har fyra bundna fettmolekyler, innebär att det kan finnas flera variabler och kombinationer av denna fosfolipid.
Syntes
Biosyntesen av kardiolipin, som man kunde förvänta sig, börjar med syntesen av fosfatidinsyra eller 1,2-diacylglycerol 3-fosfat från 3-fosfatglycerol och fettsyror och fettsyror eller fettsyror. Denna process sker i mitokondrierna i eukaryoter och i plasmamembranet av bakterier.
Kan tjäna dig: Cyklasa adenilate: egenskaper, typer, funktionerSyntes i eukaryoter
Efter bildning reagerar fosfatidinsyran med en högenergimolekyl som är analog med ATP: CTP. Då är en mellanhand, också av hög energi, känd som fosfatidyl-CMP. Den aktiverade fosfatidilgruppen överförs till hydroxylgruppen i C1 -positionen för den centrala glycerolmolekylen som fungerar som ett skelett.
Denna process resulterar. Detta får en annan fosfatidylgrupp aktiverad från en annan fosfatidyl-CMP-molekyl, en reaktion katalyserad av ett fosfatidyltransferas, även känd som kardiolipinsyntas.
Syntas -kardiolipinenzymet ligger i det inre mitokondriella membranet och verkar bilda ett stort komplex, åtminstone i jäst. Dess gen uttrycks i stora mängder i rika tyger i mitokondrier såsom hjärta, lever och skelettmuskel hos ryggradsdjur.
Regleringen av dess aktivitet beror till stor del på samma transkriptionsfaktorer och endokrina faktorer som modulerar mitokondriell biogenes.
När det är syntetiserats i det inre mitokondriella membranet måste kardiolipin omlokeras mot det yttre mitokondriella membranet så att en serie topologiska processer i membranet ges och andra strukturella element av samma rymmer rymmer.
Syntes i prokaryoter
Kardiolipininnehållet i bakterier kan vara mycket varierande och beror främst på cellernas fysiologiska tillstånd: det är vanligtvis mindre rikligt i den exponentiella fasen av tillväxt och mer rikligt när det minskar det (till exempel i den stationära fasen, till exempel).
Dess biosyntetiska väg kan skjutas av olika stressiga stimuli såsom energiunderskott eller osmotisk stress.
Fram till bildningen av fosfatidylglycerol är processen i eukaryoter och prokaryoter ekvivalent, men i prokaryoterna får fosfatidylglycerolen genom transcesserifiering en fosfatidylgrupp från en annan fosfatidylglycerolmolekyl. Denna reaktion katalyseras av ett enzymfosfolipas D, även känd som kardiolipinsyntas.
Det kan tjäna dig: SGLT (natriumglukostransportproteiner)Denna reaktion är känd som "transfatidilation" -reaktion (från engelska "Transposfatidylering "), Där en av fosfatidylglycerolerna fungerar som en givare av fosfatidilgruppen och den andra som en acceptor.
Funktioner
De fysiska egenskaperna hos kardiolipinmolekyler tillåter uppenbarligen vissa interaktioner som utför viktiga funktioner i den strukturella organisationen av membranen där de finns.
Bland dessa funktioner är diskriminering av vissa membrandomäner, interaktion eller "tvärbindning" med transmarketproteiner eller deras underdomäner, bland andra.
Tack vare sina fysikalisk -kemiska egenskaper erkänns kardiolipin som en lipid som inte bildar Bápara, men vars funktion kan vara stabilisering och "rymma" av transmembranproteiner i lipidbicapas.
Dess elektriska egenskaper ger särskilt funktioner i protonöverföringsprocesserna som äger rum i mitokondrier.
Även om celler kan överleva utan denna fosfolipid, har vissa studier fastställt att det krävs för att de optimala fungerar.
Referenser
- HARAYAMA, T., & Riezman, h. (2018). Förstå mångfalden av membranlipidkomposition. Nature Reviews Molecular Cell Biology, 19(5), 281-296.
- Luckey, M. (2008). Biologisk strukturell membran: Med biokemiska och biofysiska grunder. Cambridge University Press.
- Murray, r., Bender, D., Botham, K., Kennelly, s., Rodwell, V., & Weil, s. (2009). Harpers illustrerade biokemi (28: e upplagan.). McGraw-Hill Medical.
- Van Meer, G., Voelker, D. R., & Feigenson, g. W. (2008). Membranlipider: där de är och hur de beter sig. Naturrecensioner, 9, 112-124.
- Vance, J. OCH., & Vance, D. OCH. (2008). Biokemi av lipider, lipoproteiner och membran. I Ny förståelse av biokemi Vol. 36 (4: e upplagan.). Annars.