

Enolasstruktur, verkningsmekanism, funktioner
- 1369
- 342
- PhD. Emil Svensson
De Enolas Det är det enzym som ansvarar för att genomföra omvandlingen av D-2-fosfoglycerat (2PGA) i fosfoenolpyruvat (PEP) i glykolys och omvänd reaktion vid glukoneogenes, två metaboliska rutter som är en del av den cellulära energimetabolism.
Beslutet att katalysera denna reaktion i en eller annan riktning beror på den åtkomst som cellen till glukos har. Det vill säga behoven av att anpassa din ämnesomsättning till nedbrytning eller syntes för att få energi. Oumbärligt för att förverkliga dess vitala processer.
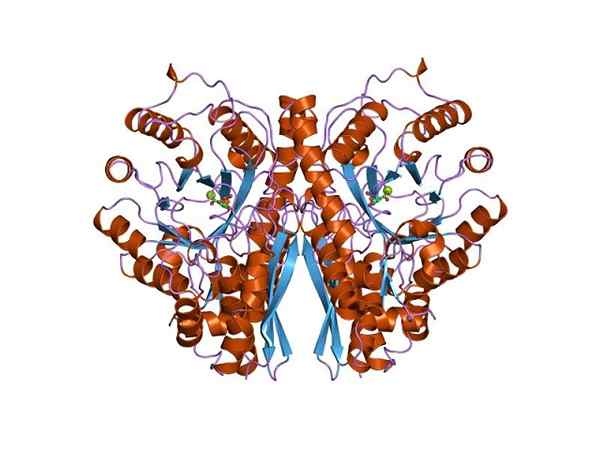
Tre -dimensionell struktur av enolasa. Av Jawahar Swaminathan och MSD -personal vid European Bioinformatics Institute [Public Domain (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons. Eftersom båda metaboliska rutter tillhör mitten av det centrala metaboliska trädet för levande varelser är det inte konstigt att aminosyrasekvensen för detta protein bevaras i arkeae, bakterier och eukaryoter. Och därför den nuvarande liknande katalytiska egenskaperna.
Platsen för enolas i cellen är begränsad till cytosol, ett fack där både glykolys (även kallad glykolys) och glukoneogenes i de flesta organismer äger rum.
Det har emellertid också detekterats i andra cellfack såsom plasmamembran av många cancerceller och celler. Där verkar det vara involverat i underlättningen av cellspridningsprocesser, en helt annan funktion än sin klassiska funktion.
Enzymer som kan utföra mer än en funktion, som enolaset gör, kallas månskenande enzymer.
[TOC]
Strukturera
Den kvartära strukturen i linus eller inte i dess ligander har bestämts i ett stort antal prokaryota och eukaryota individer.
Varje monomer presenterar två domäner: en liten aminoterminal domän och en större karboxylterminal domän. Den N-terminala domänen består av tre a och fyra ß-ark. Medan C-terminal består av åtta ß-ark som växlar mellan dem och bildar en ß-fat som är omgiven av åtta α-propeller.
Dessutom finns två unionsplatser för divalent katjoner i varje monomer som har kallats "konformationell plats" och "katalytisk plats". Den första är inte särskilt selektiv och kan ansluta sig till ett brett utbud av divalenta katjoner i frånvaro av underlag.
Kan tjäna dig: sfingolipider: vad är, egenskaper, funktioner, syntesMedan den andra binder till jonerna efter att substratet har anslutit sig till enzymet. Föreningen av joner till båda platserna är avgörande för att reaktionen ska fortsätta.
Slutligen är det viktigt att nämna att monomererna i homodimerna förenas genom att upprätthålla en parallell orientering. Därför är den aktiva platsen begränsad till den centrala regionen som bildas av nämnda union.
Men bara en av de två monomererna deltar i katalys. Detta förklarar att förmågan hos monomerer att genomföra reaktionen under experimentella förhållanden.
Handlingsmekanism
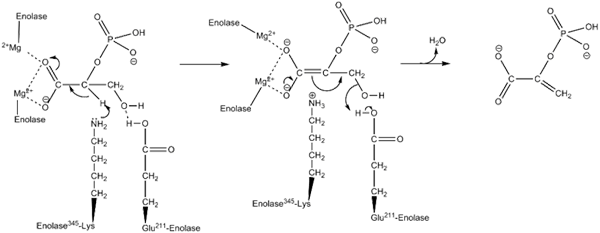 Verkningsmekanism som används av enzymet inolas. Av Kthompson08 på engelska Wikipedia [Public Domain (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons.
Verkningsmekanism som används av enzymet inolas. Av Kthompson08 på engelska Wikipedia [Public Domain (https: // CreativeCommons.Org/licenser/BY-SA/4.0)], från Wikimedia Commons. Strukturella studier, liksom de som har tillåtit att bestämma de kinetiska och fysikalisk -kemiska egenskaperna hos enolasa, har gjort det möjligt för oss att förstå deras verkningsmekanism.
Det sätt på vilket enzymet utför katalysen av reaktionen är ganska intressant. Även om ett enda underlag är involverat är en ordnad sekventiell mekanism den som har föreslagits.
Detta börjar med föreningen av en jon av Mg2+ till den konformationella platsen för en av monomererna. Kontinuerligt med föreningen mellan substratet till det aktiva stället följt av föreningen av en andra jon till det katalytiska stället och avslutas med den snabba frisättningen av produkten när reaktionen har genomförts. Vid denna tidpunkt förblir Mg2+ kopplad till den konformationella platsen.
Längs samma linjer, för att gynna förverkligandet av reaktionen det genomsnittliga enzymet i första hand genereringen av en karbanion mellanhand, eliminerar en proton av kol 2 i 2PGA. Detta gör det tack vare verkan av en grundläggande aminosyrarester.
I sekventiellt sker kol 3 hydroxylborttagning genom verkan av en sur rest från enzymet. Vid denna tidpunkt utförs föreningen av båda kolhydrater med hjälp av en dubbelbindning som bildar pep. På detta sätt kulmineras reaktionen.
Det kan tjäna dig: Monera Kingdom: Egenskaper, klassificering och exempelFunktioner
Många av de studerade enzymerna kan utföra en mängd olika funktioner som inte är relaterade till deras "klassiska funktion" i olika cellulära fack. Dessa enzymer har kallats "månsken" enzymer.
I detta avseende kan enolas betraktas som en enzymmåneshing, eftersom många funktioner som motsätter sig deras klassiska funktion har tillskrivits datum både i bakterier och eukaryoter.
Några av dessa funktioner är följande:
- Delta i underhållet av cellform och i vesikulär trafik när du interagerar med cytoskelettproteiner.
- I kärnan hos däggdjursceller fungerar som en transkriptionsfaktor som reglerar uttrycket av gener associerade med cellproliferation. Samarbeta i upprätthållandet av stabiliteten hos RNM i nedbrytningen i bakterier.
- I patogener, till exempel Streptococcus pneumoniae och Trypanosoma cruzi, Det verkar som om det fungerar som en viktig virulensfaktor.
- Det har också visat sig i Streptococcus pyogenes, Enolas utsöndras till det extracellulära medium som underlättar vävnadsnedbrytning och undvikelsen av värdsystemet.
- På ytan av tumörceller uttrycks det genom att förbättra metastaser.
Enolas och dess förhållande till cellspridningsmekanismer
Många patogener såväl som tumörceller uttrycker i sitt membran eller utsöndrar de extracellulära mediumproteaserna som kan förnedra proteinproteiner.
Denna kapacitet gör att dessa celler kan bryta igenom vävnaderna och spridas snabbt över hela värdorganismen. Därmed gynna undvikelsen av immunsystemet och därför upprättandet av infektionen.
Även när enolasa saknar proteasaktivitet, deltar i processen att sprida många patogener i dess värd såväl som tumörceller under målstastas.
Detta uppnår det tack vare det faktum att det uttrycks på ytan av dessa celler som fungerar som en plasminogenreceptor. Det senare är zimogenen i ett serinprototeas som kallas plasminat som är en del av det fibrinolytiska systemet och ACTS nedbrytande extracellulära matrisproteiner.
Därför är enolaset uttryckt på ytan en strategi som dessa celler har förvärvat för att fastställa infektionen och sprida framgångsrikt.
Det kan tjäna dig: Sympatric Speciation: Concept, Egenskaper och exempelDenna strategi består av två processer:
- Undvikande av värdens immunsystem. Sedan när dessa celler från ett värdprotein är belagt ignoreras de av immunsystemceller som känner igen icke -ägda patogener associerade med patogener.
- Spridning av plasminogen efter. Vars deltagande i nedbrytningen av extracellulära matrisproteiner, underlättar sedan den snabba och effektiva spridningen.
Referenser
- Avilan L, Gualdron-Lopez M, Quiñones W, González-González L, Hannaert V, Michels Paa, Concepción JL. Enolase: En nyckelaktör i metabolismen och en sannolik virulensfaktor för trypanosomatidparasiter-perspektiv för dess användning som ett terapeutiskt mål. Enzymforskning. 2011 Vol. Artikel ID932549, 14 sidor.
- Bhowmick I, Kumar N, Sharma S, Coppens I, Jarori GK, Plasmodium falciparum enosoase: scenspecifikt uttryck och subcellulär lokalisering. Malariajournal. 2009; 8 (1). Artikel 179.
- Dag I, Peshavaria M, Quinn GB, en differentiell molekylär klocka i enoas -isoproteinutveckling. Journal of Molecular Evolution. 1993; 36 (6): 599-601.
- av Torre-Scuder E, Manzano-Román R, Pérez-Sánchez R, Siles-Lucas M, Oleaga A. Kloning och karakterisering av plasminogenbindande yta -socierad enolaas från Schistosoma Bovis. Veterinär parasitologi. 2010; 173: 73-84.
- Dinovo EC, Boyer PD. Isotopiska sonder av enolasreaktionsmekanismen. Inledande och jämlikhetsisotope Valutakurser: Primär- och sekANDARY -isotopeffekter. J Biol Chem. 1971; 246 (14): 4586-4593.
- Kaberdin VR, Lin-Chao S, avslöjar nya roller för mindre komponenter i E. Coli RNA -nedbrytning. RNA -biologi. 2009; 6 (4): 402-405.
- Keller A, Peltzer J, Carpentier G. Interaktioner mellan enoler. Biochimica et Biophysica Acta. 2007; 1770 (6): 919-926.
- Lung J, Liu KJ, Chang JY, Leu SJ, Shih NY. MBP-1 kodas effektivt av ett alternativt transkript av den enri1-genen buttranslationellt reglerad av proteasomberoende proteinförskjutning. FEBS JOURNAL. 2010; 277 (20): 4308-4321.
- Pancholi v. Multifunktionellt a-enolas: dess roll i sjukdomar. Cellular and Molecular Life Sciences. 2001; 58 (7): 902-920.
- Poyner RR, Cleland WW, Reed GH. Roll av metallins i katalys av enolas. En ordnad kinetisk mekanism för ett enda substratenzym. Biokemi. 2001; 40: 9008-8017.
- Segovia-Gamboa NC, Chávez-Munguía B, Medina-Flores A, Entamoeba invadens, Encystationsprocess och enolas. Experimentell parasitologi. 2010; 125 (2): 63-69.
- Tanaka M, Sugisaki K, Nakashima K, Byt i nivåer av översättningsbara mRNA för enoaseisozymer under utveckling av kikn -skelettmuskel. Biokemisk och biofysisk forskningskommunikation. 1985; 133 (3): 868-872.