

Glykolysfunktioner, enzymer, faser, produkter, betydelse
- 1765
- 6
- Anders Larsson
De Glykolys O Glykolys är den huvudsakliga glukoskatabolismvägen, vars ultimata mål är att få energi i form av ATP och NADH reducerande kraft, från detta kolhydrat.
Denna rutt, helt klargjorde på 1930 -talet av Gustav Embden och Otto Meyerhof medan de studerar glukosförbrukning i skelettmuskelceller, består av fullständig oxidation av nämnda monosackarid och i sig själv representerar en anaerob väg för att erhålla energi.
Molekylstrukturen i ATP, en av glykolytiska produkterna (sammanfattning av den glykolytiska vägen (källa: Tekks på engelska Wikipedia/CC BY-SA (https: // Creativecommons.Org/licenser/BY-SA/3.0) via Wikimedia Commons) via Wikimedia Commons) Det är en av de viktigaste metaboliska rutterna, eftersom det förekommer med sina skillnader, i alla levande organismer som finns, encellulär eller multicellulär, prokaryoter eller eukaryoter, och man tror att det är en kedja av reaktioner som är mycket bevarade evolutionärt i naturen i naturen i naturen.
Det finns faktiskt vissa organismer och typer av celler som uteslutande beror på denna väg för att besöka.
I första hand består glykolys i oxidation av glukos, av 6 kolatomer, till pyruvat, som har tre kolatomer; Med den samtidiga produktionen av ATP och NADH, användbar för celler från metabolisk och syntetisk synvinkel.
I de celler som kan ytterligare bearbeta de produkter som erhållits från glukoskatabolism slutar glykolys med produktion av koldioxid och vatten genom Krebs -cykeln och elektrontransportkedjan (aerob glykolys).
Tio enzymatiska reaktioner äger rum under den glykolytiska vägen och även om regleringen av dessa reaktioner kan vara något annorlunda från en art till en annan, är regleringsmekanismerna också ganska bevarade.
[TOC]
Glykolysfunktioner
Från metabolisk synvinkel är glukos en av de viktigaste kolhydraterna för alla levande varelser.
Det är en stabil och mycket löslig molekyl, så den kan transporteras med relativt enkelhet i hela djurets eller en växt, varifrån den lagras och/eller erhålls till där den behövs som cellulärt bränsle.
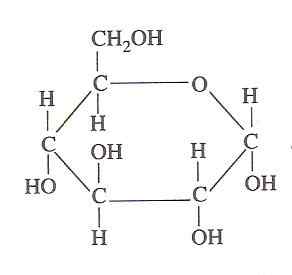 Glukosstruktur (källa: oliva93/cc by-s (https: // creativecommons.Org/licenser/BY-SA/3.0) via Wikimedia Commons)
Glukosstruktur (källa: oliva93/cc by-s (https: // creativecommons.Org/licenser/BY-SA/3.0) via Wikimedia Commons) Den kemiska energin som finns i glukos utnyttjas av levande celler genom glykolys, som består av en serie extremt kontrollerade steg genom vilka energin som frigörs från oxidationen av detta kolhydrat i mer användbara energiformer av energi kan "fångas", därmed dess betydelse.
Genom denna rutt erhålls inte bara energi (ATP) och reducerande kraft (NADH), utan den ger en serie metaboliska mellanhänder som är en del av andra rutter, också viktiga från det anabola (biosyntetiska) och den allmänna cellulära operationen. Här är en lista:
- Glukos 6-fosfat för Pentosas-fosfatvägen (PPP, engelska Pantos Mushco -väg)
- Pyruvate för mjölkfermentering
- Pyruvate för aminosyrasyntes (alanin, främst)
- Pyruvate för trikarboxylsyran
- Fruktos 6-fosfat, glukos 6-fosfat och dihydroxyacetonfosfat, som fungerar som "konstruktionsblock" på andra vägar såsom glykogensyntes, fettsyror, triglycerider, nukleotider, aminosyror, etc.
Energiproduktion
Mängden ATP som produceras av den glykolytiska vägen.
Men när det gäller aeroba celler fungerar glykolys också som en nödsituationskälla och fungerar som ett "förberedande steg" före oxidativa fosforyleringsreaktioner som karakteriserar celler med aerob metabolism.
Enzymer som deltar i glykolys
Glykolys är bara möjlig tack vare deltagandet av de 10 enzymerna som katalyserar reaktionerna som kännetecknar denna väg. Många av dessa enzymer är alosteriska och förändringsform eller konformation när de utövar sina katalytiska funktioner.
Det finns enzymer som bryter och bildar kovalenta bindningar mellan sina underlag och det finns andra som kräver att specifika kofaktorer utövar sina funktioner, främst metalljoner.
Strukturellt sett har alla glykolytiska enzymer ett centrum som väsentligen bildas av ark p -parallell omringad av Hellices a och beställde i mer än en domän. Dessutom kännetecknas dessa enzymer eftersom deras aktiva platser vanligtvis finns i unionens platser mellan domäner.
Det är också viktigt att lyfta fram att huvudregleringen av rutten går genom kontroll (hormonella eller metaboliter) av enzymer såsom hexokinas, fosfofrucerachinas, glyceraldehyd 3-fosfatdehydrogenas och pyruvatkinas.
Kan tjäna dig: Pulmonal parenkyma: Beskrivning, histologi, sjukdomar Huvudregleringspunkter för den glykolytiska vägen.Org/licenser/BY-SA/3.0) via Wikimedia Commons)
Huvudregleringspunkter för den glykolytiska vägen.Org/licenser/BY-SA/3.0) via Wikimedia Commons) 1- Hexoquinase (HK)
Den första reaktionen av glykolys (glukosfosforylering) katalyseras av hexokinas (HK), vars verkningsmekanism verkar bestå av en "inducerad justering" av substratet, som främjar "stängningen" av enzymet runt ATP och av glukos ( hans underlag) när de har anslutit sig till dessa.
Beroende på den organisme som beaktas kan det finnas en eller flera isoenzymer, vars molekylvikt sträcker sig mellan 50 (cirka 500 aminosyror) och 100 kDa, eftersom de verkar vara grupperade i form av gränser, vars bildning gynnas av den Närvaro av glukos, magnesiumjoner och ATP.
Hexokinas har en tertiär struktur bestående av alfa- och beta -ark öppna, även om det finns många strukturella skillnader i dessa enzymer.
2- Isomeras fosfoglukos (PGI)
Den fosforylerade glukosen med hexokinas isomeriseras till 6-fosfat fruktos med hjälp av isomeras fosfoglukos (PGI), även känd som glukos 6-fosfatisomeras. Enzymet tar då inte bort eller lägger till atomer, utan ordnar dem snarare på strukturnivå.
Detta är ett aktivt enzym i sin dimoriska form (monomeren väger mer eller mindre 66 kDa) och är involverat inte bara i glykolys, utan också i glukoneogenes, i syntesen av kolhydrater i växter, etc.
3- fosfofrutequinase (PFK)
6-fosfatfruktosen är substrat för fosfofrucerachinasenzymet, som kan fosforylering igen denna molekyl som använder ATP som en fosforylgruppdonator, vilket producerar 1,6-biffosfatfruktos.
Detta enzym finns i bakterier och däggdjur som ett homotrameriskt enzym (sammansatt av fyra identiska underenheter av 33 kDa vardera för bakterier och 85 kDa vardera i däggdjur) och i jäst är det en oktameter (sammansatt av större underenheter, mellan 112 och 118 kDa).
Det är ett alosteriskt enzym, vilket innebär att det är positivt eller negativt reglerat av någon av dess produkter (ADP) och andra molekyler såsom ATP och citrat.
4- Aldolas
Also known as fructose 1.6-biffosphate Aldolase, aldolase catalyzes the catalytic rupture of fructose 1.6-biphosphate in dihydroxyacetone phosphate and glyceraldehyde 3-phosphate and reaction reaction, that is, the union of both sugars for the formation of the formation of Fructose 1.6-biphosphate.
Med andra ord, detta enzym skär fruktosen 1,6 bifosfat precis i mitten och släpper två fosforylerade föreningar av 3 kolatomer. Aldolase består också av fyra identiska underenheter, var och en med sin egen aktiva webbplats.
Förekomsten av två klasser (I och II) av detta enzym har bestämts, som skiljer sig åt med mekanismen för reaktionen som katalyserar och eftersom vissa (den första) förekommer i "lägre" bakterier och eukaryoter, och de andra (de andra (de andra (de andra den andra) är i bakterier, protister och metazoos.
Aldolaset av de "överlägsna" eukaryoterna består av en homotermeter av underenheter med 40 kDa molekylvikt, var och en bildad av ett fat sammansatt av 8 p/a -ark.
5- Triosa-fosfatisomeras (TIM)
De två triosa-fosforylerade kan konverteras med varandra tack vare verkan av isomeras-triosa-fosfat, vilket gör att båda sockerarter kan användas under hela glykolysen, vilket säkerställer fullständig användning av varje glukosmolekyl som kommer in i vägen.
Detta enzym har beskrivits som det "perfekta" enzymet, eftersom det katalyserar den reaktion som beskrivs om en miljard gånger snabbare än skulle hända utan dess deltagande. Dess aktiva plats ligger i mitten av en beta-fatstruktur, ett kännetecken för många glykolytiska enzymer.
Det är ett dimérica -protein, bildat av två identiska underenheter på cirka 27 kDa, båda med en kulaformad struktur.
6- Glyceraldehyd 3-fosfatdehydrogenas (GAPDH)
3-fosfatglyceraldehyden som produceras genom verkan av isomerastriosa-fosfatet fungerar som ett substrat för GAPDH, som är ett homoterrameriskt enzym (34-38 kDa varje underenhet) som samarbetar med en NAD+ -molekyl i varje av dess 4-aktiva platser , liksom 2 fosfat- eller sulfatjoner.
I denna passage av rutten tillåter enzymet fosforylering av ett av dess substrat som används oorganiskt fosfat som en fosforylgruppsgivare, med samtidig reduktion av två NAD+ -molekyler och produktionen av 1,3-bifospphoglyceratoatoatoatoatous.
7- FOSFOGLYCERATO KINASE (PGK)
Fosfoglyceratkinas ansvarar för att överföra en av de 1,3-bifosfoglyceratfosfatgrupper till en ADP-molekyl på grund av fosforylering på underlagsnivån. Detta enzym använder en mekanism som liknar den som används av hexokinas, eftersom det stängs till kontakt på dess underlag och skyddar dem från interferensiella vattenmolekyler.
Kan tjäna dig: Aponeurosis: Egenskaper, funktioner, platsDetta enzym, som andra som använder två eller flera underlag, har en bindande plats för ADP och ett annat för fosfatsocker.
Till skillnad från de andra beskrivna enzymerna är detta protein en 44 kDa -monomer med en bilobulär struktur, sammansatt av två domäner av samma storlek anslutna med ett smalt "spår".
8- mutas fosfoglycerate
3-fosfoglyceratet genomgår en förändring från fosfatgruppen till kol 2, mitt i molekylen, som representerar en strategisk instabilitetsplats som underlättar den efterföljande överföringen av gruppen till en ATP-molekyl i den sista reaktionen av rutten.
Denna bakre reglo katalyseras av enzymet fosfoglycerat mutasa, ett dimérica -enzym för människor och tetrameriska för jäst, med en underenhetsstorlek nära 27 kDa.
9- Enolas
Enolas katalyserar uttorkning av 2-fosfoglycerat till fosfoenolpiruvat, nödvändigt steg för generering av ATP i följande reaktion.
Det är ett dimérica -enzym som består av två identiska underenheter på 45 kDa. Det beror på magnesiumjoner för stabilitet och för den konformationella förändring som krävs för att gå med i ditt underlag. Det är en av de enzymer som uttrycks med större överflöd i cytosolen hos många organismer och övningar ytterligare funktioner till glykolytik.
10- Quinasa Piruvato
Den andra fosforyleringen på substratnivån som äger rum i glykolys katalyseras av kinaspyruvatet, som är ansvarig för överföringen av fosforylgruppen av fosfoenolpiruvatet till ADP och produktion av pyruvat.
Detta enzym är mer komplex än någon av de andra glykolytiska enzymerna och hos däggdjur är det ett homotramalt enzym (57 kDa/subenhet). Det finns, i ryggradsdjur, minst 4 isoenzymer: L (i levern), R (i erytrocyter), M1 (i muskler och hjärna) och M2 (fostervävnad och vuxna vävnader).
Glykolysfaser (steg för steg)
Den glykolytiska vägen består av tio sekventiella steg och börjar med en glukosmolekyl. Under processen är glukosmolekylen "aktiverad" eller "beredd" med tillägg av två fosfater och investerar två ATP -molekyler.
Därefter är det "klippt" i två fragment och modifieras slutligen kemiskt ett par gånger, och syntetiserar, på vägen, fyra ATP -molekyler, så nettoförstärkningen motsvarar två ATP -molekyler.
Från ovanstående kan man dra slutsatsen att rutten är uppdelad i en energi "investering" -fas, grundläggande för den fullständiga oxidationen av glukosmolekylen och en annan fas av energi "förstärkning", där energin som används initialt fylls och två ATP -nät molekyler.
- Energiinvesteringsfas
1- Det första steget i den glykolytiska vägen består av fosforylering av glukos medierad av hexokinas (HK), för vilket enzymet använder en ATP-molekyl för varje glukosmolekyl som är fosforylerad. Det är en irreversibel reaktion och beror på närvaron av magnesiumjoner (Mg2+):
Glukos + ATP → Glukos 6-fosfat + ADP
2- 6-fosfatglukos som således produceras isomeriseras till 6-fosfat fruktos tack vare verkan av isomeras fosfoglukosenzym (PGI) (PGI). Detta är en reversibel reaktion och innebär inte en ytterligare energiförbrukning:
Glukos 6-fosfat → fruktos 6-fosfat
3- Därefter innebär ett annat energiinvesteringssteg fosforylering av fruktos 6-fosfat för att bilda 1,6 bifosfat fruktos. Denna reaktion katalyseras av enzymet fosfofrutquinase-1 (PFK-1). Liksom det första steget i rutten är fosfatgruppens donatormolekyl ATP och det är också en irreversibel reaktion.
Fruktos 6-fosfat + ATP → Fruktos 1.6-bifosfat + ADP
4- I detta steg av glykolys finns det katalytiska brottet av fruktos 1,6-bifosfat i dihydroxyacetonfosfat (DHAP) en ketos och i glyceraldehyd 3-fosfat (GAP) en aldosa. Denna aldoliska kondensation katalyseras av enzymet aldolas och är en reversibel process.
Fruktos 1.6-biffosfat → dihydroxyacetonfosfat + glyceraldehyd 3-fosfat
5- Den sista reaktionen av energiinvesteringsfasen består i sammankopplingen av trioas DHAP-fosfat och gap som katalyseras av triosa-fosfatisomeras (TIM) enzym, ett faktum som inte kräver ytterligare energididering och som också är en reversibel process.
Dihydroxyacetonfosfat ↔ glyceraldehyd 3-fosfat
- Energiförstärkningsfas
6- 3-fosfatglyceraldehyd används "nedströms" på den glykolytiska vägen som ett substrat för ett oxidationsreaktion och ett annat fosforylering, katalyserat av samma enzym, glykeraldehyd 3-fosfat dehydrogenas (gapdh).
Enzymet katalyserar oxidationen av kol C1 för molekylen till en karboxylsyra och fosforylering av detta i samma position, vilket producerar 1,3-bifoscicerat. Under reaktionen reduceras 2 NAD+ -molekyler per glukosmolekyl och 2 oorganiska fosfatmolekyler används.
Kan tjäna dig: luftrör2glyceraldehyd 3-fosfat + 2NAD + + 2PI → 2 (1,3-bifosfoglycerato) + 2NADH + 2H
I aeroba organismer passerar varje NADH som produceras på detta sätt genom elektrontransportkedjan för att tjäna som ett substrat för syntes av 6 ATP -molekyler på grund av oxidativ fosforylering.
7- Detta är det första syntessteget av ATP vid glykolys och innebär verkan av kinasfosfoglycerato (PGK) på 1,3-bifosfoglycerat, överföring av en fosforylgrupp (fosforylering vid substratnivån) från denna molekyl till en molekyl ADP, ger 2atp 2Atp och 2 molekyler 3-fosfoglycerat (3pg) för varje glukosmolekyl.
2 (1,3-bifosfoglycerato) + 2Adp → 2 (3-fosfoglycerate) + 2atp
8- 3-fosfoglyceratet fungerar som ett substrat för enzymet fosfoglycerato mutasa (PGM), vilket gör det 2-fosfoglycerat genom förskjutning av fosforylgruppen av kol 3 till kol 2 med ett två steg reaktion som är reversibelt och beroende av Magnesiumjoner (mg+2).
2 (3-fosfoglycerate) → 2 (2-fosfoglycerate)
9- Enzymet i ett 2-fosfoglycerat dehydrat och producerar fosfoenolpiruvat (PEP) med hjälp av en reaktion som inte förtjänar tillsats av ytterligare energi och som syftar till att producera en hög energiförening, som kan donera sin fosforylgrupp i nästa reaktion.
2 (2-fosfoglycerate) → 2fosfoenolpiruvate
10-fosfoenolpiruvat är ett substrat av pyruvatkinas (Pyk) enzymet, som är ansvarigt för överföringen av fosforylgruppen i denna molekyl mot en ADP-molekyl och därigenom katalysering av fosforylering vid underlaget nivå i nivå.
I reaktionen produceras 2atp och 2 pyruvatmolekyler för varje glukos och närvaron av kalium och magnesium i jonisk form är nödvändig.
2FOSPO ELPLIVATO + 2ADP → 2Pyruvate + 2Atp
Nettoprestationen för glykolys består på detta sätt av 2Atp och 2NAD+ för varje glukosmolekyl som kommer in i rutten.
Om dessa är celler med aerob metabolism, producerar den totala nedbrytningen av en glukosmolekyl mellan 30 och 32 ATP genom Krebs -cykeln och elektrontransportkedjan.
Glykolysprodukter
Den allmänna reaktionen av glykolys är som följer:
Glukos + 2NAD + + 2Adp + 2Pi → 2Pyruvate + 2atp + 2NADH + 2H+
Därför, om det kort analyseras, kan det säkerställa att de viktigaste produkterna från den glykolytiska vägen är pyruvat, ATP, NADH och H.
Men den metaboliska destinationen för varje reaktionsförmedlare beror, i stor utsträckning, på cellulära behov, varför alla mellanhänder kan betraktas som reaktionsprodukter, att kunna mynta dem enligt följande:
- Glukos 6-fosfat
- Fruktos 6-fosfat
- Fruktos 1,6 bifosfat
- Dihydroxyacetonfosfat och glyceraldehyd 3-fosfat
- 1.3-bifosfoglycerato
- 3-fosfoglycerate och 2-fosfoglycerat
- Fosfoenolpiruvat och pyruvat
Betydelse
Även om glykolys, i sig själv (kunde prata om anaerob glykolys) producerar endast cirka 5% av ATP som kan extraheras från aerob katabolism av glukos, är denna metaboliska väg grundläggande av flera skäl:
- Det fungerar som en "snabb" energikälla, särskilt i situationer där ett djur måste lämna ett viltillstånd snabbt, för vilket aerob oxidationsprocesser inte skulle vara tillräckligt snabba.
- Skelett "vita" muskelfibrer i människokroppen, till exempel, är snabba sammandragningsfibrer och beror på anaerob glykolys för att fungera.
- När en cell av någon anledning behöver glykolytisk.
- Många celler är beroende av glukos som en energikälla efter glykolytisk väg, inklusive röda blodceller, saknar inre organeller och ögonceller (särskilt de i hornhinnan) som inte har en stor mitokondria densitet.
Referenser
- Canback, b., Andersson, s. G. OCH., & Kurland, c. G. (2002). Den globala fylogeni av glykolytiska enzymer. Proceedings of the National Academy of Sciences, 99 (9), 6097-6102.
- Chaudhry R, Varacallo M. Biokemi, glykolys. [Uppdaterad 2019 AP 21]. I: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2020 jan-. Tillgänglig från: https: // www.Ncbi.Nlm.Nih.Gov/böcker/nbk482303/
- Fothergill-Gilmore, L. TILL., & Michels, s. TILL. (1993). Utveckling av glykolys. Framsteg inom biofysik och molekylärbiologi, 59 (2), 105-235.
- Kim, j. W., & Dang, c. V. (2005). Mångfacetterade roller för glykolytiska enzymer. Trender inom biokemiska vetenskaper, 30 (3), 142-150.
- Kumari, a. (2017). Sweet Biochemistry: Att komma ihåg strukturer, cykler och vägar av Mnemonics. Akademisk press.
- Li, x. B., Gu, j. D., & Zhou, Q. H. (2015). Granskning av aerob glykolys och dess viktiga enzymer-nya mål för lungcancerterapi. Thoracic Cancer, 6 (1), 17-24.