

Kärnporegenskaper, funktioner, komponenter
- 644
- 44
- Johan Eriksson
han kärnpore (Från det grekiska, porer = steg eller transit) är "dörren" till kärnan som tillåter det mesta av transporten mellan nukleoplasma och cytoplasma. Kärnporen går med i de inre och yttre membranen i kärnan för att bilda kanaler, som tjänar för transport av proteiner och RNA.
Ordet Poro återspeglar inte komplexiteten i strukturen i fråga. På grund av detta är det att föredra att hänvisa till Nuclear Pore Complex (CPN), istället för kärnkraftspore. CPN kan uppleva förändringar i sin struktur under transport eller tillstånd i cellcykeln.
Nyligen har det upptäckts att nukleoporiner, proteiner som utgör CPN, har en viktig roll för att reglera genuttryck. Således, när mutationer inträffar som påverkar funktionen hos nukleoporiner, produceras patologier hos människor, såsom autoimmuna, kardiomyopatier, virala infektioner och cancer.
[TOC]
Egenskaper
Genom elektrontomografi fastställdes att CPN har en tjocklek av 50 nm, en yttre diameter mellan 80 och 120 nm och en inre diameter på 40 nm. Stora molekyler såsom den stora underenheten av ribosomer (PM 1590 kDa) kan exporteras utanför kärnan genom CPN. Det uppskattas att det finns mellan 2000 och 4000 cpn per kärna.
Molekylvikten för en individuell CPN är ungefär mellan 120 och 125 MDA (1 MDA = 106 Da) i ryggradsdjur. Däremot är CPN mindre i jäst, där den har cirka 60 MDA. Trots den enorma storleken på CPN är nukleoporiner mycket bevarade i alla eukaryoter.
Translokalisering genom CPN är en snabb process, vars hastighet är 1000 translokationer/andra. CPN bestämmer emellertid inte själva transportflödets riktning.
Detta beror på RANGTP -gradienten, som är större i kärnan än i cytosolen. Denna lutning upprätthålls av en guaninbytarfaktor sprang.
Under cellcykeln upplever CPNS montering och oenighetscykler. Monteringen sker vid gränssnittet och omedelbart efter mitos.
Funktioner
Ribonukleinsyra (liten nukleär RNA, messenger -RNA, överföring av RNA), proteiner och ribonukleoproteiner (RNP) måste aktivt transporteras genom CPN. Detta innebär att energin från ATP och GTP -hydrolys krävs. Varje molekyl transporteras på ett specifikt sätt.
I allmänhet är RNA -molekyler packade med protein som bildar RNP -komplex och exporteras på detta sätt. Däremot måste proteiner som aktivt transporteras till kärnan ha ett sekvenstecken på plats i kärnan (SLN), innehavare av aminosyravfall med positiv belastning (till exempel kKKRK).
Kan tjäna dig: mesosomProteiner som exporteras till kärnan måste ha en exportsignal till kärnan (NES) som är rik på leucinaminosyra.
Förutom att underlätta transport mellan kärnan och cytoplasma är CPN: er involverade i kromatinorganisation, genuttrycksreglering och DNA -reparation. Nukleoporiner (NUP: er) främjar transkriptionsaktivering eller förtryck, oavsett cellproliferationstillstånd.
I jäst finns nuptarna i CNP för kärnkraftsförpackningen. I metazoarierna är de inne. De utför samma funktioner i alla eukaryoter.
Import av ämnen
Genom CPN finns en passiv spridning av små molekyler i både riktningar och aktiv transport, proteinimport, export av RNA och ribonukleoproteiner (RNP) och den dubbelriktade molekylerna Shuttle. Det senare inkluderar RNA, RNP och proteiner involverade i signalering, biogenes och ersättning.
Importen av proteiner i kärnan sker i två steg: 1) proteinbindning till den cytoplasmiska sidan av CPN; 2) ATP -beroende translokalisering genom CPN. Denna process behöver ATP -hydrolys och utbyte av GTP/BNP mellan kärnan och cytoplasma.
Enligt en transportmodell rör sig det mottagande proteinkomplexet längs kanalen genom unionen, dissociationen och unionen igen till de upprepade FG-sekvenserna av nukleoporinerna. På detta sätt rör sig komplexet från en nukleoporin till en annan inuti CPN.
Export av ämnen
Liknar import. RAN GTPASE sätter riktning mot transport genom CNP. RAN är en molekylväxel med två konformationella tillstånd, beroende på om den är kopplad till BNP eller GTP.
Två specifika reglerande proteiner utlöser omvandlingen mellan de två tillstånden: 1) cytosolisk GTPas-aktiverande protein (GAP), som producerar GTP-hydrolys och därmed konverterar RAN-GTP till RAN-BNP; och 2) Nuclear Guanina Exchange Factor (GEF), som främjar utbytet av BNP av GTP och konverterar RAN-BNP till RAND-GTP.
Cytosol innehåller huvudsakligen RAN-BNP. Kärnan innehåller huvudsakligen Ran-GTP. Denna lutning av de två konformationella formerna av RAN dirigerar transport i lämplig adress.
Importen av mottagaren, tillsammans med positionen, underlättas av unionen till repetitionerna-FG. Om du når kärnkraftssidan av CNP går Ran-GTP med mottagaren för att släppa din position. Således skapar RAN-GTP riktningen för importprocessen.
Kärnkraftsexport är liknande. Ran-GTP i kärnan främjar emellertid föreningen av positionen till exportmottagaren. När exportmottagaren rör sig genom poren mot cytosolen möter den RAN-GAP, vilket inducerar GTP-hydrolys till BNP. Slutligen frigörs mottagaren från sin position och Ran-BNP i cytosolen.
Kan tjäna dig: polymorfonukleära leukocyterRNA -transport
Exporten av vissa slags RNA liknar exporten av proteiner. Till exempel använder ARNT och RNANS (liten kärnkraft) RANGTP -lutningen och transporteras genom CPN med hjälp av medel. Exporten av mogna ribosomer är också beroende av RANGTP -lutningen.
MRNA exporteras på ett helt annat sätt än proteinerna och andra RNA. För sin export bildar mRNA ett RNP -messenger (RNPM) -komplex, där en RNM -molekyl är omgiven av hundratals proteinmolekyler. Dessa proteiner har bearbetningen, Tätning, Empalme och polyadentilly av MNA.
Cellen måste kunna skilja mellan RNPM med mogen RNM och RNPM med omoget mRNA. MRNA, som bildar RPNM -komplexet, kan anta topologier som måste ombyggas för transport. Innan RNPM kommer in i CPN inträffar ett kontrollsteg som utförs av tramp- och exosomproteinkomplex.
När mogen RNPM monteras transporteras RPNM genom kanalen med hjälp av en transportmottagare (NXF1-NXT1). Denna mottagare behöver ATP -hydrolys (inte en RANGTP -lutning) för att fastställa riktningens ombyggnad av RNPM, som kommer att nå cytoplasma.
Kärnporkomplexet och kontrollen av genuttryck
Vissa studier indikerar att CPN -komponenter kan påverka regleringen av genetiskt uttryck genom kontroll av kromatinstruktur och dess tillgänglighet till transkriptionsfaktorer.
I eukaryoter av nyare utveckling är heterokromatin företrädesvis beläget på periferin av kärnan. Detta territorium avbryts av Euchromatin -kanaler, som upprätthålls av CPN -kärnkorgen. Kärnkorgföreningen med euchromatin är relaterat till gentranskription.
Exempelvis innebär aktivering av transkription på CPN -nivån interaktionen mellan komponenterna i kärnkorgen med proteiner såsom histonacetyltransferas -saga och RNA -exportfaktorer.
Således är kärnkorgen en plattform för många underhållsgener (Hushållning) starkt transkriberade och gener inducerade starkt av förändringar i miljöförhållanden.
Kärnkrafts- och virologikomplexet
Viral eukaryotcellinfektion beror på CPN. I varje fall av virusinfektion beror dess framgång på DNA, RNA eller RPN som passerar genom CPN för att uppnå sitt slutliga mål, vilket är replikationen av viruset.
Kan tjäna dig: cytoplasmiska inneslutningar: vad är, egenskaper, funktionerAPE -virus 40 (SV40) har varit en av de mest studerade modellerna för att undersöka CPN: s funktion vid translokalisering i kärnan. Detta beror på att SV40 har ett litet genom (5.000 baser).
Det har visats att transporten av virus -DNA underlättas proteinproteiner från viruset, som skyddar viruset tills kärnan når.
Komponenter
CPN är inbäddad i kärnhöljet och består av mellan cirka 500 och 1000 nupt. Dessa proteiner är organiserade i strukturella underkomplex eller moduler, som interagerar med varandra.
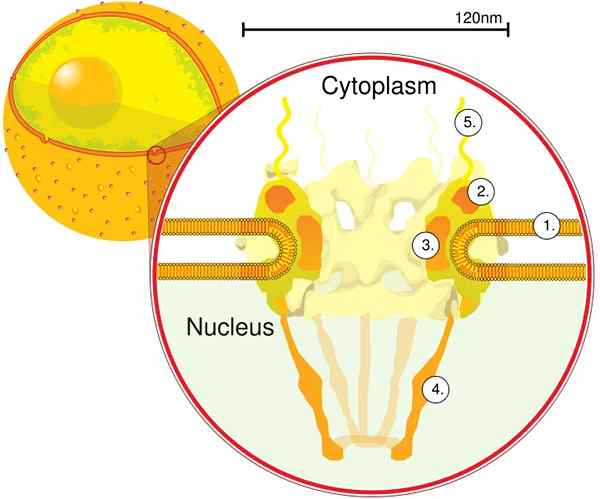
Den första modulen är en central komponent, eller ring, inuti poren i form av en sandklocka, som är begränsad av ytterligare 120 nm diameter på båda sidor, intranukleära och cytoplasmiska. Den andra modulen är kärnan och cytoplasmaringarna (120 nm i diameter vardera) som ligger runt den sandwatformade komponenten.
Den tredje modulen är åtta filament som projiceras från 120 nm -ringen inom nukleoplasma och bildar en korgformad struktur. Den fjärde modulen består av filamenten som projiceras till sidan av cytoplasma.
Det Y -formade komplexet, bestående av sex Nups och SEH 1 och Sec 13 -proteiner, är det största och bäst karakteriserade komplexet i CNP. Detta komplex är den väsentliga enheten som är en del av CPN -ställningen.
Trots den låga likheten mellan NUPS -sekvenserna är CPN -ställningen mycket bevarad i alla eukaryoter.
Referenser
- Beck, m., Skada, e. 2016. Kärnkraftskomplexet: Förstå dess funktion genom strukturell insikt. Nature Reviews, Molecular Cell Biology, DOI: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Kärnporproteiner och kontroll av genomfunktioner. Gener och utveckling, 29, 337-349.
- Kabachinski, g., Schwartz, T.ELLER. 2015. Kärnkraftskomplexet - struktur och funktion på ett överblick. Journal of Cell Science, 128, 423-429.
- Knocknhauer, K.OCH., Schwartz, T.ELLER. 2016. Kärnkraftskomplexet som flexibel och dynamisk grind. Cell, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.W. 2014. Den multifunktionella kärnkraften efter komplex: en plattform för att kontrollera genuttryck. Nuvarande åsikt om cellbiologi, doi: 10.1016/j.Ceb.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Republikation: Kärnkraftskomplexet - En omfattande översyn av struktur och funktion. International Journal of Academic Medicine, 3, 51-59.
- Tran, E.J., Wente, s.R. 2006. Dynamisk kärnkraft efter komplex: Livet på kanten. Cell, 125, 1041-1053.