

Succinato dehydrogenasstruktur, funktion, reglering, sjukdomar
- 1267
- 11
- Per Eriksson
SUccinato dehydrogenas (Sdh), även känd som komplexet II i elektrontransportkedjan, är det ett mitokondriellt proteinkomplex med enzymatisk aktivitet som fungerar både i Krebs -cykeln och i elektrontransportkedjan (cellandning).
Det är ett enzym som finns i alla aeroba celler. I eukaryoter är det en komplex som är nära förknippad med det inre mitokondriella membranet, medan det i prokaryoterna finns i plasmamembranet.
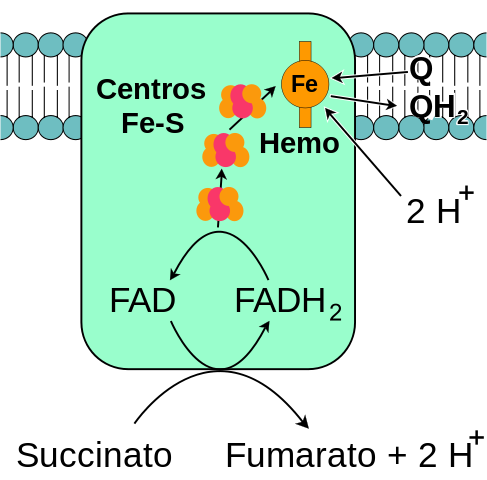 Allmänt schema för det mitokondriella komplexet succinatdehydrogenas (källa: jag själv, baserat på fvasconcellos -vektorisering. / Public Domain, via Wikimedia Commons)
Allmänt schema för det mitokondriella komplexet succinatdehydrogenas (källa: jag själv, baserat på fvasconcellos -vektorisering. / Public Domain, via Wikimedia Commons) Den komplexa succinatdehydrogenasen, upptäckt omkring 1910 och renades för första gången 1954 av Singer och Kearney, har studerats omfattande av flera skäl:
- Det fungerar både i Krebs -cykeln (citronsyrcykel eller trikarboxylsyrcykel) och i elektrontransportkedjan (katalyserar oxidationen av succinat till fumarat)
- Dess aktivitet regleras av olika aktivatorer och hämmare och
- Det är ett komplex associerat med: järn som inte är kopplat till en hemo -grupp, labylsvavel och dyucleotider av Flavina Adenina (FAD)
Det är kodat av kärngenomet och det har bevisats att mutationer i de fyra generna som kodifierar var och en av dess underenheter (a, b, c och d) resulterar i olika kliniska bilder, det vill säga de kan vara ganska negativa från punkten synen på människors fysiska integritet.
[TOC]
Strukturera
Det enzymkomplexsuccinatdehydrogenas bildas av fyra underenheter (heterotarámero) kodat av kärngenomet, så det är det enda komplexet av oxidativ fosforylering i elektrontransportkedjan som inte har någon underenhet kodad av det mitokondriella genomet.
Dessutom är detta komplex det enda som inte pumpar protoner genom det inre mitokondriella membranet under dess katalytiska verkan.
Enligt studier baserade på det enzymatiska komplexet av svinhjärtceller består den komplexa succinatdehydrogenaset av:
- en "huvud" Hydrofilisk som sträcker sig från det inre mitokondriella membranet till mitokondriell matris och
- en "linje" Hydrofob som är inbäddat i det inre mitokondriella membranet och som har ett litet segment som projiceras mot det lösliga intermembranutrymmet i mitokondrierna
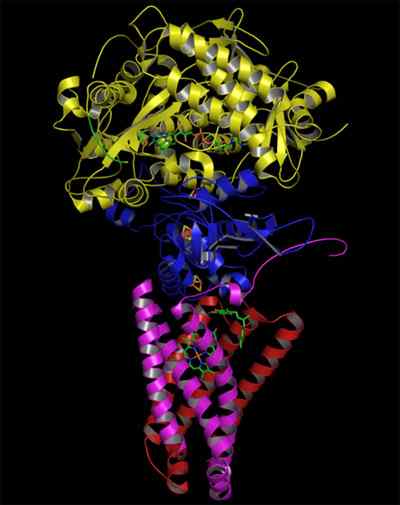
 Struktur av den komplexa succinatdehydrogenaset (källa: Zephyris på det engelska språket Wikipedia/CC BY-SA (http: // Creativecommons.Org/licenser/BY-SA/3.0/) via Wikimedia Commons)
Struktur av den komplexa succinatdehydrogenaset (källa: Zephyris på det engelska språket Wikipedia/CC BY-SA (http: // Creativecommons.Org/licenser/BY-SA/3.0/) via Wikimedia Commons) Hydrofil delstruktur
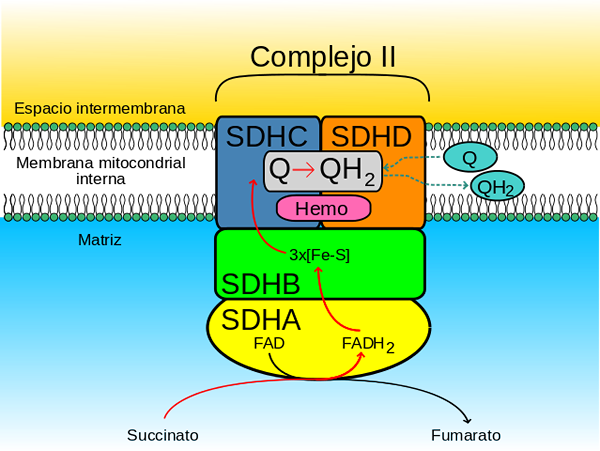
Det hydrofila huvudet består av SDHA (70 kDa) och SDHB (27 kDa) (SDH1 och SDH2 i jäst) och detta innefattar det katalytiska centrumet för komplexet.
SDHA- och SDHB -underenheter innehåller redoxkofaktorer som deltar i överföringen av elektroner till ubikinonet (Coenzyme Q10, en molekyl som transporterar elektroner mellan andningskomplex I, II och III).
SDHA-underenheten har en moofaktor (ett koenzym som deltar i oxidreduceringsreaktioner) tillsammans med dess struktur, precis på platsen för korsningen för succinat (huvudsubstratet för enzymet).
SDHB-underenheten har 3 järn-sockercentra (FE-S) som förmedlar överföringen av elektroner till ubiquinon. Ett av centren, 2Fe-2s, ligger nära SDHA-underenheten och de andra (4FE-4S och 3FE-4s) ligger intill den första.
Kan tjäna dig: fylogeniDet bör noteras att strukturstudier indikerar att SDHB -underenhet bildar gränssnittet mellan den hydrofila katalytiska domänen och domänen för "ankare" till membranet (hydrofob) i komplexet.
Hydrofob delstruktur
Komplexets membrandomän, som sagt, består av SDHC (15 kDa) och SDHD (12-13 kDa) (SDH3 och SDH4 i jäst), som är omfattande membranproteiner bildade, vardera, av 3 transmembranala propellers.
Denna domän innehåller en hemo -del b gick med i gränssnittet mellan SDHC- och SDHD -underenheterna, där var och en av de två histidinligander som håller dem tillsammans.
I detta enzym har två unionsplatser för Ubiquinona upptäckts: en av stor affinitet och en av låg affinitet.
Webbplatsen med hög affinitet, känd som Qp (P av proximal) Det är ansikte mot mitokondriell matris och bildas av specifika aminosyrarester belägna i SDHB, SDHC och SDHD.
Låga affinitetsplatsen, även kallad QD (D av distal) Det är, i den del av det inre mitokondriella membranet där komplexet sätts in, närmare intermembranutrymmet, det vill säga längre från organellmatrisen.
Tillsammans har det totala komplexet en molekylvikt nära 200 kDa och det har fastställts att det har ett förhållande på 4.2-5.0 flavin nanomoles per milligram protein och 2-4 g järn för varje mol av flavina.
Fungera
Det enzymatiska sucinatkomplexet dehydrogenas uppfyller en viktig funktion i mitokondrierna, eftersom inte bara deltar i Krebs -cykeln (där den deltar i nedbrytningen av acetyl -CoA), utan också en del av andningskedjan, grundläggande för energiproduktion ATP -formad.
Med andra ord är det ett viktigt enzym för mellanhandsmetabolism och ATP Aerob produktion.
- Det ansvarar för oxidationen av sucipen till fumarat i citronsyran Cycle
- Den matar komplexet III i elektrontransportörskedjan med elektronerna härrörande från oxidationen av sucipen, vilket hjälper till att minska syre och bilda vatten
- Elektrontransport genererar en elektrokemisk lutning genom det inre mitokondriella membranet, som gynnar ATP -syntes
Som ett alternativ kan elektroner användas för att minska molekyler från en ubiquinonas "pool", vilket ger nödvändiga reducerare för att minska superoxidanjoner som härstammar från samma andningskedja eller som kommer från exogena källor.
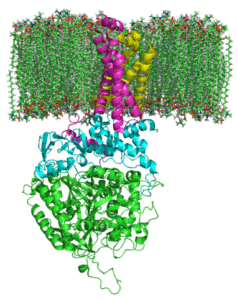
 Succate Complex Degidrogenasa (källa: Johnhfst / Public Domain, via Wikimedia Commons)
Succate Complex Degidrogenasa (källa: Johnhfst / Public Domain, via Wikimedia Commons) Hur agerar det?
Underenheten A i komplexet (som är kovalent kopplat till FAD -koenzymet) ansluter sig till substrat, fumarat och succinat, liksom dess fysiologiska regulatorer, oxalacetat (konkurrerande hämmare) och ATP.
ATP förskjuter unionen mellan oxalacetat och SDH -komplexet och sedan överförs elektronerna som är "förbi" från succinatet till SDHA -underenheten till grupperna av järn- och svavelatomer närvarande i SDHB -underenheten med hjälp av koenzymfaden.
Kan tjäna dig: myosin: egenskaper, struktur, typer och funktionFrån underenhet B når dessa elektroner hemosidorna b av SDHC- och SDHD.
Det elektroniska flödet från succinatet genom dessa transportörer och till och med den slutliga acceptorn, som är syre, är kopplad till syntesen av 1.5 ATP -molekyler för varje elektroniskt vridmoment genom andningskedjefosforylering.
Inzima defekter
Det har rapporterats att mutationer i genen som kodar för underenheten A i den komplexa succinatdehydrogenaset kan orsaka encefalopatier under barndomen, under tiden mutationerna i generna som kodar underenheter B, C och D har associerats med bildning av tumörer.
Reglering
Aktiviteten för den komplexa succinatdehydrogenaset kan regleras av post -translationella modifieringar såsom fosforylering och den acetylering, Även om aktiv platsinhibering också kan uppstå.
Acetylering av viss lysinavfall kan minska aktiviteten hos detta enzym och denna process utförs av ett enzym av acetylas känt som SIRT3; Fosforylering har samma effekt på enzymet.
Utöver dessa modifieringar regleras SDH -komplexet också av mellanhänder av Krebs -cykeln, särskilt Oxalacetat och den suberda. Oxalacetat är en kraftfull hämmare, medan succinat gynnar dissociationen av oxalacetat, fungerar som aktivator.
Impulcinatbrist dehydrogenas
Succinatbrist dehydrogenas är en anomali eller mitokondriell andningskedjestörning. Denna brist orsakas av SDHA (eller SDHAF1), SDHB, SDHC och SDHD.
Olika undersökningar har visat homozygota och heterozygota mutationer i dessa gener, särskilt SDHA. Mutationerna av dessa gener orsakar aminosyrasubstitutioner i proteinet (i någon av SDHA, B, C eller D) eller misslyckas som kodifierar onormalt korta proteiner.
Därför ger aminosyrasubstitutioner och onormalt kort proteinkodning upphov till störningar eller förändringar av SDH -enzymet, vilket orsakar ett fel i den optimala kapaciteten hos mitokondrier att producera energi. Detta är vad forskare kallar som mitokondriell andningskedjestörning.
Denna störning kan uttryckas hos människan fenotypiskt på många sätt. De mest kända är: brist eller brist på språklig utveckling, spastisk quadruplex, muskelinvilliga sammandragningar (dystoni), muskelsvaghet och myokardiopati, bland andra relaterade problem.
Vissa patienter med succinatbrist dehydrogenas kan manifestera Lights sjukdom eller Kearns-Saire-syndrom.
Hur upptäcks dehydrogenerad succinatbrist?
Vissa studier tyder på användning av kvalitativa tester och analys, såväl som kvantitativ, enzymatisk, andningskedja biokemisk analys. Andra, å andra sidan, föreslår den fullständiga expansionen med polymeraskedjereaktionen (PCR) av exonerna av underenheterna som studeras och sedan respektive sekvensering.
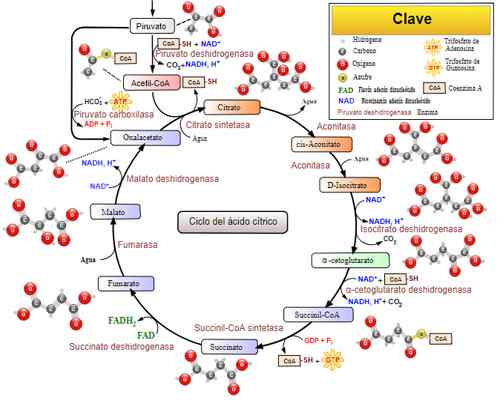 Tricarboxylsyrcykel (Krebs -cykel). Tagen och redigerad från: Narayanese, Wikiuserpedia, Yassinemrabet, Totobaggins (översatt till spanska av Alejandro Porto) [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)].
Tricarboxylsyrcykel (Krebs -cykel). Tagen och redigerad från: Narayanese, Wikiuserpedia, Yassinemrabet, Totobaggins (översatt till spanska av Alejandro Porto) [CC BY-SA 3.0 (https: // CreativeCommons.Org/licenser/BY-SA/3.0)]. Relaterade sjukdomar
Det finns många fenotypiska uttryck som produceras av mitokondriella andningskedjor, på grund av succinatbristdehydrogenas. Men när det handlar om syndrom eller sjukdomar är det samtal om följande.
Kan tjäna dig: de 8 viktigaste biogeokemiska cyklerna (beskrivning)Avlärningssyndrom
Det är en progressiv neurologisk sjukdom, associerad med mutationer i kärngenomet (i detta fall av dehydrogenas succinat), som påverkar pyruvat-diaktydrogenaskomplexet till den oxidativa fosforyleringsvägen.
Symtomen dyker upp före individens första år, men i atypiska fall har de första symtomen under tonåren observerats.
Bland de mest observerade symtomen är: hypotoni med förlust av cefalisk kontroll, ofrivilliga rörelser, återkommande kräkningar, andningsproblem, oförmåga att flytta de okulära blodkropparna, pyramiden och extrapiramidala tecken bland andra. Anfall är inte särskilt vanliga.
Det är möjligt att sjukdomen kan upptäckas i prenatala diagnoser. Ett specifikt botemedel eller behandling är inte känt, men vissa specialister föreslår behandlingar med vissa vitamin eller kofaktorer.
Gastrointestinal stroma tumör (GIST)
Vanligtvis kallad GIST, det är en typ av gastrointestinal Tract -tumör, som i allmänhet utvecklas i områden som mage eller tunntarmen. Det tros att orsaken till dessa beror på en viss grupp högspecialiserade celler som kallas ICC -celler eller interstitiella celler i Cajal.
Andra överväganden om orsaken till GIST är mutationerna av vissa typer av gener, som enligt vissa författare orsakar 90% av tumörerna. Generna är: kit, pdgfra -gener, dehydrogenas succinat (SDH) - dålig.
Succinatdehydrogenas (SDH) - Dålig förekommer främst hos unga kvinnor, producerar magtumörer och med relativ frekvens producerar metastaser i lymfkörtlar. En liten procentandel förekommer hos barn och i de flesta fall beror det på bristen på uttryck för SDHB -underenheten.
Kearns-Sayre-syndrom
Det har fastställts att vissa patienter med succinatdehydrogenasbrister kan manifestera Kearns-Sayre-syndromet. Denna sjukdom är relaterad till mitokondriella störningar och kännetecknas av frånvaron av rörelse av okulära ballonger.
Andra egenskaper hos denna sjukdom är pigmentär retinit, dövhet, kardiomyopati och centrala nervsystemets tillstånd. Vanligtvis observeras dessa symtom innan patienten fyller 20 år. Ingen prenatal diagnos för detta tillstånd är känt.
Botemedel mot denna sjukdom. Behandlingen är palliativ, det vill säga det fungerar bara för att minska effekterna av sjukdomen, inte botemedel. Å andra sidan, även om det beror på antalet drabbade organ och den mottagna medicinska vården, är livslängden relativt normal.
Referenser
- Ackrell, b. TILL., Kearney, E. B., & Singer, T. P. (1978). [47] däggdjursuccinatdehydrogenas. I metoder i enzymologi (Vol. 53, sid. 466-483). Akademisk press.
- Brère, J. J., Favier, J., Ghouzzi, v. OCH., Djoudi, f., Benit, s., Gimenez, a. P., & Rustin, s. (2005). Succinat dehydrogenasdefikicy hos människa. Cellular and Molecular Life Sciences CMLS, 62 (19-20), 2317-2324.
- Cecchini, g., Schröder, jag., Gunalus, r. P., & Maklashina och. (2002). Succinat dehydrogenas och uppfyller redaktas från Escherichia coli. Biochimica et Biophysica Acta (BBA) -Bioenergetics, 1553 (1-2), 140-157.
- Hatefi och., & Davis, K. TILL. (1971). Succinat dehydrogenas. Yo. Rening, molekylära egenskaper och understruktur. Biochemistry, 10 (13), 2509-2516.
- Hedersstedt, l. TILL. R. S., & Rutberg, L. TILL. R. S. (1981). Succinat dehydrogenas-en jämförande översyn. Mikrobiologiska recensioner, 45 (4), 542.
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger principer för biokemi. Macmillan.
- Rutter, J., Winge, d. R., & Schiffman, J. D. (2010). Succinat dehydrogenas-membly, reglering och roll i mänsklig diew. Mithochondrion, 10 (4), 393-401.
- « Väsentliga fettsyror Funktioner, betydelse, nomenklatur, exempel
- Regel t -funktioner, så att det tjänar, exempel »