

Tre egenskaper och funktioner i organismen
- 1920
- 340
- Anders Larsson
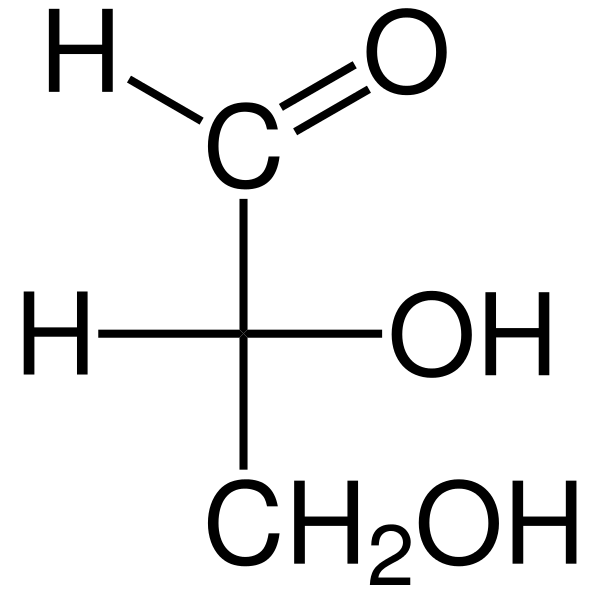
De Trios De är tre -kolmonosackarider vars empiriska kemiska formel är c3H6ANTINGEN6. Det finns två trios: glyceraldehyd. Triosas är viktiga i metabolismen eftersom de förbinder tre metaboliska vägar: glykolys, glukoneogenes och pentosfosfatvägen.
Under fotosyntesen är Calvin-cykeln en källa till trios som tjänar för fruktos-6-fosfatbiosyntes. Detta socker, på fosforylerat sätt, omvandlas av enzymatiskt katalyserade steg till reserv eller strukturella polysackarider.
Källa: Wesalius [Public Domain] Trioserna deltar i biosyntesen av lipider som ingår i cellmembran och adipocyter.
[TOC]
Egenskaper
Glyceraldehyden Aldosa har en quiral kolatom och därför har den två enantiomeros, L-glyceraldehyden och D-glyceraldehyden. Både enantiomerer D och L har olika kemiska och fysiska egenskaper.
D-glyceraldehyden bröt det polariserade ljusets plan till höger (+) och har en rotation [a] D, vid 25 ° C, från +8.7 °, medan L-glyceraldehyden bröt planet för det polariserade ljuset till vänster (-) och har en rotation [a] D, vid 25 ° C, från -8.7 °.
Det chirala kolet av glyceraldehyd är kol 2 (C-2), vilket är en sekundär alkohol. Fischers projektion representerar hydroxylgruppen (-OH) för D-glyceraldehyden till höger och OH-gruppen i vänster L-Glyce.
Dihydroxyaceton saknar chirala kol och har inte enantiomeriska former. Tillsatsen av en hydroximetylengrupp (-CHOH) till glyceraldehyd eller dihydroxyaceton gör det möjligt att skapa ett nytt chiralcenter. Följaktligen är socker en tetrosa eftersom det har fyra kolhydrater.
Tillägget av en grupp -chah till Terosa skapar ett nytt chiralt centrum. Bildat socker är en pentos. Grupper -Chah kan fortsätta att läggas till högst tio kol kol.
Kan tjäna dig: fylogeniFunktioner i organismen
Triosas som mellanhänder i glykolys, glukoneogenes och pentosa -fosfatvägen
Glykolys består av brottet av glukosmolekylen i två pyruvatmolekyler för att producera energi. Denna rutt innebär två faser: 1) förberedande fas eller energiförbrukning; 2) Energiproduktionsfas. Den första är den som producerar trioserna.
I den första fasen ökas glukosfri energiinnehåll genom bildning av fosfoésters. I denna fas är adenosintrofosfatet (ATP) fosfatdonatorn. Denna fas kulminerar med omvandlingen av fruktos fosfoéster 1,6-bifosfat (F1.6BP) i två trioasfosfat, glyceraldehyd 3-fosfat (GA3P) och dihydroxyaceton-fosfat (DHAP).
Glukoneogenes är glukosbiosyntes från pyruvat och andra mellanhänder. Använd alla glykolysenzymer som katalyserar reaktioner vars biokemiska standardenergivariation är i jämvikt (ΔGº '~ 0). På grund av detta har glykolys och glukoneogenes vanliga mellanhänder, inklusive GA3P och DHAP.
Pentosfosfatvägen består av två steg: en oxidativ fas av glukos-6-fosfat och en annan bildning av NADPH och ribosa-5-fosfat. I den andra fasen omvandlas 5-fosfatribosen till glykolysförmedlare, F1.6BP och GA3P.
Triosa och Calvin -cykeln
Fotosyntes är uppdelad i två steg. I den första inträffar lätta beroende reaktioner som producerar NADPH och ATP. Dessa ämnen används i den andra, där det finns fixering av koldioxid och hexosbildning från trios genom en väg som kallas Calvin -cykel.
I Calvin-cykeln katalyserar det 1,5-bififera-oxygenas (Rubisco) ribulosenzymet den kovalenta föreningen av Co2 Till pentos ribulosa 1,5-bifosfat och bryter den instabila mellanhand av sex kolatomer i två molekyler av tre kolatomer: 3-fosfoglyceratet.
Kan tjäna dig: oligosackarider: egenskaper, komposition, funktioner, typerGenom enzymatiska reaktioner som inkluderar fosforylering och reduktion av 3-fosfoglycerate, med användning av ATP och NADP, inträffar GA3P. Denna metabolit omvandlas till 1,6 bifosfat fruktos (F1,6bp) med en metabolisk väg som liknar glukoneogenes.
Genom verkan av ett fosfatas omvandlas F1,6bp till fruktos-6-fosfat. Sedan producerar en isomeasfosfhexos glukos 6-fosfat (GLC6P). Slutligen omvandlar en epikeras GLC6P till glukos 1-fosfat, som tjänar till stärkelsebiosyntes.
Trios och lipider i biologiska och adipocytmembran
GA3P och DHAP kan bilda fosfatglycerol som är en nödvändig metabolit för biosyntes av triacylglyceroler och glycerolipider. Detta beror på att båda trioasfosfat kan konverteras av en reaktion katalyserad av triosa -isomerasfosfatet, som håller båda trioserna i jämvikt.
Glycerol-fosfat-enzymet dehydrogenas katalyserar en oxidreduktionsreaktion, där NADH donerar ett par elektroner till DHAP för att bilda 3-fosfatglycerol och NAD+. L-glycerol 3-fosfat är en del av skelettet av fosfolipider som är en strukturell del av biologiska membran.
Glycerolen är proquiral, den saknar asymmetriska kol, men när en av dess två primära alkoholer bildar en fosfoester kan den korrekt kallas L-glycerol 3-fosfat eller D-glycerol 3-fosfat.
Glyceofosfolipider kallas också fosfoglycerider, som utses till fosfatidinsyra härledd. Fosfoglycerider kan bilda fosfoacylglyceroler genom att bilda esterbindningar med två fettsyror. I detta fall är den resulterande produkten 1,2-fosfodiacylglycerol, vilket är en viktig del av membranen.
En glyceofosfas katalyserar hydrolysen av 3-fosfatglycerolfosfatgruppen, vilket ger glycerol mer fosfat. Glycerol kan tjäna som en startmetabolit för biosyntes av triacylglycerider, som är vanliga i adipocyter.
Kan tjäna dig: koagulasetest: grund, procedur och användningArkeobakteriens triosas och membran
I likhet med eubakterier och eukaryoter bildas 3-fosfatglycerol från trioasfosfat (GA3P och DHAP). Det finns emellertid skillnader: den första är att 3-fosfatglycerol i arkeobakteriemembran är L-konfiguration, medan i eubakterier och eukaryota membran är det konfiguration D.
En andra skillnad är att arkeobakterimembran bildar ester kopplingar till två långa kolvätekedjor av isoprenoidgrupper, medan i eubakterier och eukaryoter glycerol bildar esterbindningar (1,2-diacilglycerol) med två kolvätekedjor av fettsyror.
En tredje skillnad är att i arkeobakteriemembran skiljer sig ersättare i fosfat och 3-fosfatglycerol från Eubacteria och Eukaryotes. Till exempel är fosfatgruppen kopplad till disackarid a-Glukopiranso- (1®2)-p-Galatofuranosa.
Referenser
- Cui, s. W. 2005. Matkolhydrater: Kemi, fysiska egenskaper och applikationer. CRC Press, Boca Raton.
- Kuk, s., Mäkinen, K, Honkala och., Saag, m., Kennepohl, E., Eapen, a. 2016. Erythritol är mer effektivt än xylitol och sorbitol vid hantering av orala hälsoändpunkter. International Journal of Dentistry.
- Nelson, D. L., Cox, m. M. 2017. Lehninger principer för biokemi. W. H. Freeman, New York.
- Sinnott, m. L. 2007. Kolhydratkemi och biokemi struktur och mekanism. Royal Society of Chemistry, Cambridge.
- Stick, r. V., Williams, s. J. 2009. Kolhydrater: livets väsentliga molekyler. Elsevier, Amsterdam.
- Voet, D., Voet, j. G., Pratt, C. W. 2008. Fundamentals of Biochemistry - Life på molekylnivå. Wiley, Hoboken.
- « Aldosegenskaper, typer, antal kolhydrater
- Fluoxetin (prozac) mekanism för verkan och biverkningar »