

Archaea -domän

- 4315
- 962
- Karl Johansson
 Archaeasillustration
Archaeasillustration Vad är archaea -domänen?
han Archaea -domän o Archaea Kingdom representerar en av de tre livets domäner. Det består av mycket liknande mikroskopiska encelliga organismer och samtidigt mycket annorlunda än bakterier och eukaryoter i många aspekter.
Förekomsten av denna grupp demonstrerades nyligen, mer eller mindre i slutet av 1970 -bågarna, även kallad arkeobakterier.
Woee- och kollaboratörsstudier baserades på jämförande fylogenetiska analyser mellan sekvenserna av ribosomala RNA för alla levande varelser, och begreppet tre domäner upprätthålls, trots den enorma mängden sekvenser som fortsätter att läggas till kataloger av ribosomalt RNA (databaser).
Dessa studier lyckades skymta möjligheten att bågarna är en brodergrupp av eukaryoterna, trots deras likheter med sina prokaryoter (bakterier), så att de kan representera den "förlorade länken" mellan prokaryoter och eukaryoter.
Det antas att dessa mikroorganismer representerar mer än 20% av alla befintliga prokaryoter i oceaniska vatten, mellan 1 och 5% i jord och den mest dominerande gruppen i marina sediment och geotermiska livsmiljöer.
Bågarna kännetecknas av att bebor under ”extrema” förhållanden som varma källor, salinor, miljöer med mycket höga temperaturer och mycket surt pH, ogästvänliga platser där syrekoncentrationen är mycket reducerad eller noll, etc.
Evolutionärt ursprung och rFylogenetiska elvor
Det finns tre teorier om det möjliga ursprunget till de tre livets domäner som föreslagits av Woese och kollaboratörer:
- Bakterier divergerade först och bildade en avstamning som producerade bågar och eukaryoter
- En "Proto-Eucariotic" -linje divergerade från en helt probotisk avstamning (den för bakterier och bågar)
- Bågarna divergerade av en avstamning som senare gav upphov till eukaryoter och bakterier
1989 föreslog två forskare, Gogarten och Iwabe, självständigt ett sätt att fylogenetiskt analysera alla levande organismer (en omöjlig sak att göra från studierna av unika gensekvenser).
Iwabe använde analysen av sekvenserna av gener som producerats i "tidiga" händelser av genduplikation, "rota" livets träd från jämförelsen av sekvensen för papegojgenerna som kodar för töjningsfaktorer.
Töjningsfaktorer är GTP -bindande proteiner som deltar i översättningen, specifikt i föreningen av överföring RNA -molekyler amioacilerade till ribosomer och i translokationen av peptidilöverföringen.
Enligt resultaten från jämförelserna mellan sekvenserna för de tre grupperna är generna som kodar för töjningsfaktorerna i bågarna mer lik de hos eukaryota organismer än de av bakterier.
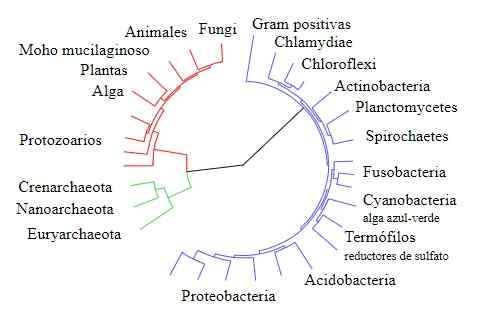 Livets träd
Livets träd Gogarten jämförde å andra sidan sekvenserna av andra gener som produceras av dupliceringshändelser, särskilt de som kodar för typ V och typ F -underenheter av ATPasenzymet som finns i bågar/eukaryoter respektive i bakterier.
Resultaten som erhållits av Gogarten, som de som anges ovan, visar att dessa gener i bågarna (som påstås komma från genetiska dupliceringshändelser) är närmare besläktade med eukaryoter än med deras bakteriekoftor.
Dessa argument stöds också av analysen som genomfördes av andra forskare många år senare, som använde sekvenser från en annan familj av duplicerade gener, den för aminoacilsyntesöverföring, vilket stärkte uppfattningen av "närheten" mellan bågar och eukaryoter.
Det kan tjäna dig: Selenito -buljong: Vad är, grund, förberedelse, användningDet woese trädet
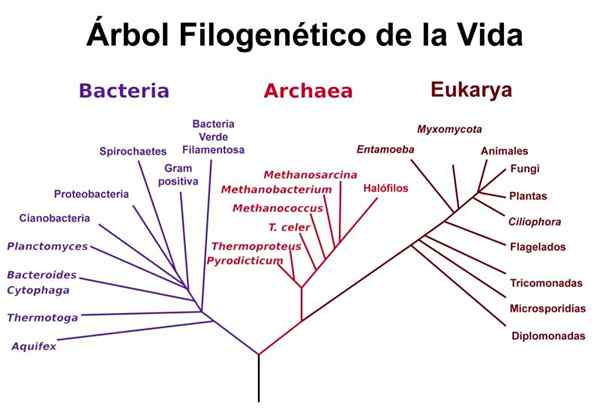 Woeses livsträd
Woeses livsträd Woese använde analyserna som utfördes av Gogarten och iwabe och andra studier relaterade till ribosomala RNA -sekvenser för att föreslå dess "version" av livets träd, där det är uppenbart att bågar och eukaryoter utgör "bröder" -grupper, separerade från bakterierna till och med Även om de ribosomala RNA -sekvenserna mellan bågar och bakterier liknar varandra mer.
Archaea domänfunktioner
Bågarna är kända för några mycket speciella (egna) egenskaper och även för att presentera "kombinationer" av egenskaper som en gång trodde var exklusiva för bakterier eller för eukaryota organismer.
- Som bakterier, bågar De är prokaryota byråer, Det vill säga, inuti är det genetiska materialet inte låst av ett membran (de har ingen kärna) och det finns inga cytoplasmiska membraniska organeller.
- De är i allmänhet mikroorganismer som liknar de för bakterier, deras DNA är i form av en Cirkulär kromosom och några mindre cirkulära fragment kända som Plasmider.
- De delar med bakterier närvaron av Topoisomeras- och turerenzymer Liknande, som representerar "indirekt" bevis på "närheten" i termer av den kromosomala strukturen hos båda grupper av organismer.
- Archaeas -gener visar emellertid en stor Homologi med många eukaryoter, särskilt med de vars upptäckt härstammar från studier med antibiotika.
- Maskinen i replikation, transkription och översättning av eukaryoter och bågar är mycket lika, särskilt med avseende på enzymet ADN -polymeras.
- Dess kodande gener för protein De har inga introner (Men andra gör), i motsats till eukaryota gener. Dessutom har bågar proteiner som liknar histoner associerade med deras DNA, närvarande i eukaryoter och frånvarande i bakterier.
- De kännetecknas av närvaron av Eterlipider av isoprenil i sina cellmembran, såväl som från frånvaron av acylsoler lipider och ett fettsyrasyntesas.
- En av underenheterna i dess RNA -polymerasenzym är uppdelat och dess messenger RNA, liksom i bakterier, har inte "capuchas" (från engelska kille) I dess ändar 5 '.
- De har ett mycket specifikt utbud av antibiotikaskänslighet och har Begränsning av typ II mycket lik de som beskrivs för bakterier.
- En annan viktig funktion har att göra med det faktum att mycket av bågarna har cellvägg, Men till skillnad från bakterier består detta inte av peptidoglycan.
Egenskaper hos dess membranlipider
Archaean membranlipider skiljer sig avsevärt från de som finns i bakterier och eukaryota organismer, och detta har betraktats som en mycket viktig differentiell egenskap.
Den huvudsakliga skillnaden mellan dessa amfipatiska molekyler (med en hydrofil polär ände och en hydrofob apolär) är att föreningen mellan den del av glycerol och kedjor av fettsyror i bågarnas lipider är genom en eterlänk, medan i bakterierna och eukaryoter motsvarar en esterlänk.
En annan viktig skillnad är att bågarna har lipider med fettsyror som kännetecknas av närvaron av mycket grenade isoprenilkedjor med metylgrupper, under tiden har eukaryoter och bakterier främst fettsyror av icke -förgrenade kedjor.
Lipiderna av eukaryoter och bakterier "konstrueras" på ett glycerolskelett till vilka fettsyrakedjorna förestras i positionerna som motsvarar kolatomer 1 och 2, men i bågarna innehåller glyceroletrar syror syror i positionerna 2 och 3.
Kan tjäna dig: Flora och Fauna of Río Negro: Representativa arterEn annan skillnad med avseende på membranlipider har att göra med dess biosyntetiska väg, eftersom vissa enzymer också är olika i bågarna.
Till exempel har vissa arter av bågar en bifunktionell överföringsprenilenzym, som är ansvarig för att tillhandahålla föregångare både för syntesen av escualen och för syntesen av glyceril-lipid isaprensoids. I bakterier och eukaryoter utförs dessa funktioner av separata enzymer.
Klassificering av bågar
Enligt uppgifterna om sekvenserna för de små underenheterna i bågarnas ribosomaler har denna grupp huvudsakligen uppdelats i två "kanter", som är kända som kanten Crenarchaeota och kanten Euryarchaeta, vars medlemmar är framför allt odlade bågar In vitro.
Många av de bågar som beskrivs nyligen har emellertid inte odlats In vitro Och de är bara avlägsna relaterade till de sekvenser som har isolerats från arten som upprätthålls i laboratorierna.
Kant Crenarchaeota
 Termofila bakterier, crenarchaeota kant
Termofila bakterier, crenarchaeota kant Denna grupp består huvudsakligen av arter av hypertermofila och termakidofila bågar, det vill säga de kön av bågar som bor i invånliga miljöer med termiska förhållanden och extremt pH.
Det består av en enda taxonomisk klass, som kallas Themoprotei, Inom vilka är de kommande fem taxonomiska orderna: Acidilobales, Desulfurokock, Glöd, Sulfolobal och Termokrotealer.
Exempel på några av genrerna som tillhör dessa klasser kan vara genrerna Sulfolobus, Desulforokock, Pyrodiktium, Themoproteus och Themofilum.
Kant Euryarchaeta
 Methanosarcina Barkeri Fusaro, Euryarchaota Filo
Methanosarcina Barkeri Fusaro, Euryarchaota Filo Medlemmarna i denna grupp har ett något bredare ekologiskt intervall, att kunna hitta någon hypertermofil, metenogen, halofil och till och med termofil mätare, nimidala bågar, svavelreducerare, järnoxidader och vissa organrofiler och vissa organotrofiler.
Taxonomiska klasser som beskrivs för euriarqueotas är åtta och är kända som Metanopyri, Metanokocker, Metanobakterier, Metanomycrobia, Arkaeglobi, Halobakterier, Termokocker och Themoplasmata.
Många av de bågar som tillhör denna grupp är i stor utsträckning distribuerade och är i jordar, sediment och marina vatten, liksom i de extrema miljöerna som beskrivs.
Kant Thaumarchaeota
 Axinella polypoider, thaumarchaeota filo. Källa: Liné1, CC BY-SA 3.0, via Wikimedia Commons
Axinella polypoider, thaumarchaeota filo. Källa: Liné1, CC BY-SA 3.0, via Wikimedia Commons Denna kant definierades relativt nyligen och mycket få av de arter som tillhör den har odlats In vitro, Så lite är känt om dessa organismer.
Alla kantmedlemmar får sin energi från ammoniumoxidation och distribueras globalt i färskvattenkroppar, jord, sediment och varma källor.
Filos Korachaeota, Aigachaeota och Garchaeota
 Korachaeota
Korachaeota Vissa expertforskare inom området, baserat på analysen av genomiska sekvenser, har nyligen bestämt förekomsten av ytterligare tre kanter i archaea -riket, även om de arter som föreslogs för dessa kanter ännu inte var isolerade i ett laboratorium.
I allmänhet har medlemmar som tillhör dessa kanter hittats under ytan av många land- och marina ekosystem, men också i termiska vatten och i djupt hav hydrotermiska system.
Näring
Majoriteten av archaean -arter med chimiórophos -organismer, det vill säga kan använda mycket reducerade oorganiska föreningar för att få den energi de behöver för att "flytta" sina metaboliska maskiner, särskilt den som har att göra med andning.
"Specificiteten" av de oorganiska molekylerna som de använder som substrat för att producera energi beror på miljön där varje speciell art utvecklas.
Andra bågar, såväl som växter, alger, briofyter och cyanobakterier, kan utföra fotosyntes, det vill säga de använder och omvandlar ljusenergin från solstrålar till användbar kemisk energi.
Det kan tjäna dig: proteinaminosyrorDet har visats att vissa bågar lever i magen (vommen) av vissa idisslare djur (bland vilka är kor, får, getter, etc.), så dessa beskrivs som "ömsesidiga bågar", eftersom de konsumerar en del av fibern som dessa djur äter och samarbetar med matsmältningen av några av deras komponenter.
Fortplantning
Precis som bakterier är bågar encelliga organismer vars reproduktion uteslutande är asexuell. De viktigaste mekanismerna som beskrivs från arten upprätthålls In vitro är:
- Binär klyvning, där varje arkea är "del" i hälften för att komma från två identiska celler
- Gemination eller "fragmentering", där celler avger "fragment" eller "delar" av sig själva som kan bilda nya, genetiskt identiska celler.
Livsmiljö
Bågarna har huvudsakligen relaterade till "extrema" miljöer, det vill säga de naturliga platser som sätter allvarliga begränsningar för normal utveckling av levande varelser, särskilt när det gäller temperatur, pH, salthalt, anaerobios (frånvaro av syre), etc.; Anledning till varför din studie är oerhört intressant, eftersom de har unika anpassningar.
De senaste teknikerna för molekylär analys för identifiering av arter av otroliga mikroorganismer (isolerade och underhållna In vitro I ett laboratorium) har de tillåtit att upptäcka närvaron av båge.
De flesta bågar som har identifierats i naturen klassificeras emellertid enligt livsmiljön de upptar, som är släktingar i litteraturen termerna "hypertermofiler", "acidofiler" och "extrema termakidofiler", "extrema halofiler" och "metanogenos".
Miljöerna som ockuperas av hypertermofilbågar är de som kännetecknas av mycket höga konstant temperaturer (långt över de "normala" temperaturerna som de flesta levande varelser utsätts för).
Miljöerna där extrema acidofiler lever, å andra sidan, är de där pH är mycket lågt och dessa kan också särskiljas av höga temperaturer (extrema termakidofiler), under tiden är miljöerna med extrema halofiler de där koncentrationen av salter är mycket hög.
Metanogena bågar lever i frånvaro av syre eller anaerobios, i miljöer där andra molekyler såsom elektronacceptorer i deras metabolism kan användas och kan producera metan som en produkt av metaboliskt "avfall.".
Exempel på bågarter
Det finns många kända arter av bågar, men här kommer bara några av dem att göras.
Ignicoccus sjukhus och Nanoarchaeum equitans
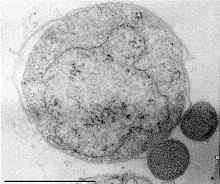 Ignicoccus sjukhus
Ignicoccus sjukhus Yo. Sjukhus Det tillhör släktet av crenarqueotas som kallas Ignicoccus och är en quimiolithotrofisk organisme som använder molekylärt väte som elektrondonator för svavelreduktion. Denna art har det minsta genomet av alla bågar som beskrivs in vitro hittills.
Yo. Sjukhus Det uppträder som en "parasit" eller "symbiote" av en annan art: Nanoarchaeum equitans. Det senare har inte odlats In vitro Och dess genom är det minsta av alla obestämt bågar som har beskrivits.
Det bor främst i marina miljöer och har inte gener för biosyntes av lipider, aminosyror, nukleotider eller kofaktorer, så experimentella bevis tyder på att det erhåller dessa molekyler tack vare dess interaktion med Yo. Sjukhus.
Sackarovoraner
Detta är en slags anaerob termakidofil arked. Det hittades för första gången i kroppar av Terrestrial Hot Springs i Kamchatka.
Stafylothermus hellenicus
Denna archea tillhör kanten av crenarqueotas, särskilt till Order of the Desulfurococcal. Det är en hypertermofil heterotrofisk Archea (bor i mycket heta miljöer) och kräver svavel för att få energi.