

Begränsningsenzymer funktioner, typer och exempel
- 3699
- 91
- Per Karlsson
De restriktionsenzymer De är endonukleaser som används av vissa bågar och bakterier för att hämma eller "begränsa" förökningen av virus inuti. De är särskilt vanliga i bakterier och är en del av deras utländska DNA -försvarssystem som kallas restriktions-/modifieringssystemet.
Dessa enzymer katalyserar det dubbla bandet -DNA på specifika platser, reproducerbara och utan användning av ytterligare energi. De flesta kräver närvaro av kofaktorer som magnesium eller andra divalenta katjoner, även om vissa också kräver ATP- eller S-adenosylmetionin.
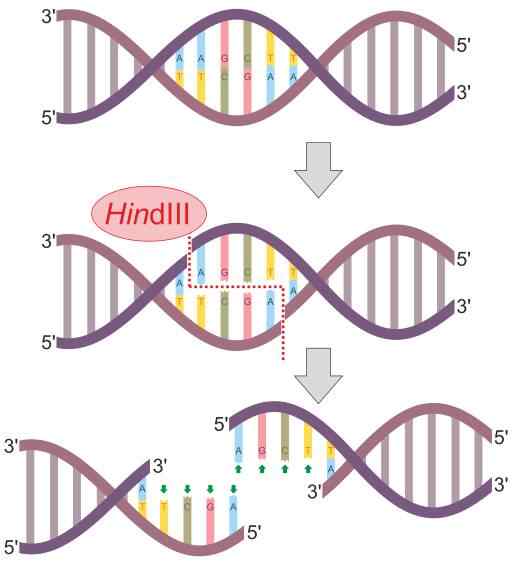
Hindii-restriktionsenzymreaktionsschema (källa: Helixitta [CC BY-SA 4.0 (https: // CreativeCommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons) Begränsningens endonukleaser upptäcktes 1978 av Daniel Nathans, Arber Werner och Hamilton Smith, som fick Nobelpriset i medicin för deras upptäckt. Hans namn härstammar generellt från organismen där de först observeras.
Sådana enzymer används allmänt i utvecklingen av DNA -kloningsmetoder och annan molekylärbiologi och genetiska teknikstrategier. Dess egenskaper för igenkänning av specifika sekvenser och skärkapacitet för sekvenserna nära igenkänningsställena gör dem kraftfulla verktyg i genetisk experiment.
Fragmenten som genereras av restriktionsenzymerna som har agerat på en viss DNA -molekyl kan användas för att återskapa en "karta" av den ursprungliga molekylen genom att använda information på de platser där enzymet skär DNA.
Vissa begränsningsenzymer kan ha samma igenkänningsställe i DNA, men de klipper inte nödvändigtvis på samma sätt. Således finns det enzymer som gör nedskärningar som lämnar romoser och enzymer som klipper och lämnar sammanhängande ytterligheter, som har olika tillämpningar inom molekylärbiologi.
Det finns för närvarande hundratals olika restriktionsenzymer kommersiellt tillgängliga, erbjuds av olika kommersiella hus; Dessa enzymer fungerar som "skräddarsydda" molekylär sax för olika ändamål.
[TOC]
Funktioner
Begränsningsenzymer uppfyller den motsatta funktionen av polymeraser, eftersom de är hydrolyserade eller bryter esterbindningen inom fosfodiésterbindningen mellan angränsande nukleotider i en nukleotidkedja.
Inom molekylärbiologi och genteknik är de allmänt använda verktyg för konstruktion av uttryck och kloningvektorer, liksom för identifiering av specifika sekvenser. De är också användbara för konstruktion av rekombinanta genom och har stor bioteknisk potential.
De senaste framstegen inom genterapi använder aktuell restriktionsenzymer för införandet av gener bestämda i vektorer som är fordon för transport av sådana gener till levande celler, och att de förmodligen har förmågan att sätta in sig i cellgenomet för att utföra permanenta förändringar.
Handlingsmekanism
Begränsningsenzymer kan katalysera DNA -DNA för dubbelband, även om vissa kan känna igen enkla band -DNA -sekvenser och till och med RNA. Snittet sker efter erkännandet av sekvenserna.
Kan tjäna dig: glioxylatcykel: egenskaper, reaktioner, reglering, funktionerVerkningsmekanismen består av hydrolysen av fosfodiésterlänken mellan en fosfatgrupp och en deoxyribos i skelettet för varje DNA av DNA. Många av enzymerna kan skära på samma plats som de känner igen, medan andra skär mellan 5 och 9 baspar före eller efter samma.
Normalt skär dessa enzymer vid 5' -änden av fosfatgruppen, vilket ger upphov till DNA -fragment med en 5' -fosforyländ och en hydroxyländ 3 'terminal.
Eftersom proteiner inte kommer i direktkontakt med DNA -igenkänningsstället måste dessa translokeras på varandra följande tider tills den specifika platsen uppnås, kanske genom "glidande" mekanismer på DNA -strängen.
Under det enzymatiska snittet placeras fosfodiésterlänken för var och en av DNA -strängarna inom en av de aktiva platserna för restriktionsenzymer. När enzymet lämnar erkännande och skärplats gör det det genom icke -specifika övergående föreningar.
Grabbar
För närvarande är fem typer av begränsningsenzymer kända. Därefter en kort beskrivning av var och en:
Begränsning av typ I
Dessa enzymer är stora pentameriska proteiner med tre underenheter, en av begränsningar, en metylering och en annan för erkännande av DNA -sekvenser. Dessa endonukleaser är multifunktionella proteiner som kan katalysera begränsningar och modifieringsreaktioner, har ATPASA och även DNA -topoisomera -aktivitet.
Enzymerna av denna typ var de första endonukleaserna som upptäcktes, de renades för första gången på 1960 -talet och sedan dess har de studerats med stort djup.
Enzymer av typ I används inte i stor utsträckning som ett bioteknologiskt verktyg, eftersom skärplatsen kan vara på ett variabelt avstånd på upp till 1.000 baspar angående erkännandewebbplatsen, vilket gör dem opålitliga när det gäller experimentell reproducerbarhet.
Begränsning av typ II
De är enzymer som består av homodimerer eller tetramrar som skär DNA i platser definierade mellan 4 och 8 bp längd. Dessa skärplatser är vanligtvis palindromiska, det vill säga de känner igen sekvenser som läses på samma sätt i båda riktningarna.
Många av typ II -restriktionsenzymer i bakterier klippte DNA när de känner igen sin främmande karaktär, eftersom det inte har de typiska modifieringarna som det egna DNA bör ha.
Dessa är enklare restriktionsenzymer eftersom de inte kräver någon annan kofaktor än magnesium (Mg+) för att känna igen och klippa DNA -sekvenserna.
Precisionen av typ II -begränsningsenzymer vid igenkänning och skärning av enkla sekvenser i DNA i exakta positioner gör dem till en av de mest använda och oumbärliga i de flesta grenar av molekylbiologi.
Kan tjäna dig: Mutualism: Egenskaper, typer, exempelInom gruppen av typ II -restriktionsenzymer klassificeras flera underklasser enligt vissa egenskaper som är unika för varje. Klassificeringen av dessa enzymer utförs genom att lägga till bokstäver i alfabetet, från A till Z efter namnet på enzymet.
Några av de mest kända underklasserna för deras användbarhet är:
Underklass IIA
De är olika underenheter Dímeros. De känner igen asymmetriska sekvenser och används som idealiska föregångare för att klippa enzymer.
Underklass IIB
De består av ytterligare en dimerer och klipper DNA på båda sidor av igenkänningssekvensen. De klippte båda DNA -strängarna i ett basparintervall senare från igenkänningsstället.
IIC underklass
Enzymer av denna typ är polypeptider med uppdelning och modifieringsfunktioner hos DNA -strängar. Dessa enzymer skär båda trådarna asymmetriskt.
Underklass IIE
Enzymerna i denna underklass är de mest använda inom genteknik. De har en katalytisk plats och kräver i allmänhet en alestroeffektor. Dessa enzymer måste interagera med två kopior av sin igenkänningssekvens för att göra en effektiv skärning. Inom denna underklass finns ECORII och ECORI -enzymer.
Begränsningsenzymer av typ III
Endonukleaser av typ III är endast sammansatta av två underenheter, den ena är ansvarig för erkännande och modifiering av DNA, medan den andra är ansvarig för nedskärningen av sekvensen.
Dessa enzymer kräver två kofaktorer för sin operation: ATP och magnesium. Begränsningsenzymerna av denna typ har två asymmetriska igenkänningsställen, translokerar DNA på ett ATP -beroende sätt och skär det mellan 20 till 30 bp intill igenkänningsstället.
Begränsning av typ IV
Enzymer av typ IV är enkla att identifiera eftersom de skär DNA med metyleringsmärken, de består av flera olika underenheter som är ansvariga för att känna igen och klippa DNA -sekvensen. Dessa enzymer använder GTP -kofaktorer och divalent magnesium.
Specifika skärplatser inkluderar nukleotidkedjor med metylerade eller hydroxymetierade cytosinrester i en eller båda trådarna av nukleinsyror.
Begränsningsenzymer av typ V
Denna klassificeringsgrupper Crisper-Cas-typenzymer, som identifierar och skär specifika DNA-sekvenser av invaderande organismer. CAS -enzymer använder en RNA -strängsyntetiserad guide för Crisper för att känna igen och attackera invaderande organismer.
Enzymer klassificerade som typ V är polypeptider strukturerade av typ I, II och II -enzymer. De kan klippa DNA -sektioner av nästan alla organisme och med ett stort antal längd. Dess flexibilitet och enkel sysselsättning gör dessa enzymer till ett av de mest använda verktygen inom genteknik för närvarande tillsammans med typ II -enzymer.
Kan tjäna dig: propionibacteriumExempel
Begränsningsenzymer har använts för detektion av DNA -polymorfismer, särskilt i populationsgenetikstudier och evolutionära studier med användning av mitokondriellt DNA för att få information om nukleotidsubstitutionshastigheter.
För närvarande har vektorerna som används för omvandling av bakterier med olika ändamål multiklonage -platser där igenkänningsställen för flera restriktionsenzymer hittas.
Bland de enzymer de mest populära är EcoRi, II, III, IV och V, erhållna och beskrivna för första gången från OCH. coli; Hindiii, från H. influensa och Bamhi av B. Amyloliquefaciens.
Referenser
- Bikle, T. TILL., & Kruger, D. H. (1993). DNA -begränsning. Mikrobiologiska recensioner, 57(2), 434-450.
- Boyaval, s., Moineau, s., Romero, D. TILL., & Horvath, s. (2007). CRISPR tillhandahåller förvärv mot virus i prokaryoter. Vetenskap, 315(Mars), 1709-1713.
- Goodsell, D. (2002). Det molekylära perspektivet: Begränsning av endonukleaser. Stamceller grundläggande i cancermedicin, tjugo, 190-191.
- Halford, s. OCH. (2001). Hopping, hoppning och slingande av restriktionsenzymer. Biokemiska samhälle transaktioner, 29, 363-373.
- Jeltsch, a. (2003). Underhåll av artens identitet och kontroll av specifikation av bakterier: En ny funktion för begränsning/modifieringssystem? Gen, 317, 13-16.
- Krebs, j., Goldstein, E., & Kilpatrick, s. (2018). Lewins gener xii (12 ed.). Burlington, Massachusetts: Jones & Bartlett Learning.
- Li och., Bröd, s., Zhang och., Ren, m., Feng, m., Peng, n.,... hon, Q. (2015). Utnyttjande typ I och typ III CRISPR-CAS-system för genomredigering. Nukleinsyror forskning, 1-12.
- Loenen, w. TILL. M., Dryden, D. T. F., Raleight, e. TILL., & Wilson, g. G. (2013). Begränsning av typ I och deras släktingar. Nukleinsyror forskning, 1-25.
- Nathans, D., & Smith, h. ANTINGEN. (1975). Begränsning av endonukleaser i analysen och omstruktureringen av DNA -molekyler. Annu. Varv. Biokemi., 273-293.
- Nei, m., & Tajima, f. (1981). DNA -polymorfism som detekteras genom restriktionsendonukleaser. Genetik, 145-163.
- Pingoud, a., Fuxreiter, m., Pingoud, v., & Wende, w. (2005). Cellular and Molecular Life Sciences Typ II Begränsning Endonukleaser: Struktur och mekanism. CMLS Cellular and Molecular Life Sciences, 62, 685-707.
- Roberts, r. (2005). Hur restriktionsenzymer blev arbetshästarna i molekylärbiologi. PNA, 102(17), 5905-5908.
- Roberts, r. J., & Murray, K. (1976). Restriktionsendonukleaser. Kritiska recensioner inom biokemi, (November), 123-164.
- Stoddard, b. L. (2005). Homing endonukleasstruktur och funktion. Kvartalsvis recensioner av biofysik, 1-47.
- Tock, m. R., & Dryden, D. T. F. (2005). Biologin för begränsning och anti-restriktion. Nuvarande åsikt i mikrobiologi, 8, 466-472. https: // doi.org/10.1016/j.Min B.2005.06.003
- Wilson, G. G., & Murray, n. OCH. (1991). Begränsnings- och modifieringssystem. Annu. Varv. Genet., 25, 585-627.
- Wu, z., & Mou, k. (2016). Genomisk insikt i Campylobacter jejuni virulens och befolkningsgenetik. Infec. Dis. Översätt. Med., 2(3), 109-19.
- Yuan, r. (1981). Struktur och mekanism för multifunktionella restriktionsendonukleaser. Annu. Varv. Biokemi., femtio, 285-315.