

Syntes av fettsyror där det händer, enzymer, steg och reaktioner
- 1431
- 79
- Anders Svensson
De fettsyrasyntes Det är processen genom vilken de grundläggande komponenterna i cellernas viktigaste lipider (fettsyror), som deltar i många mycket relevanta cellulära funktioner produceras.
Fettsyror är alifatiska molekyler, det vill säga de är i huvudsak sammansatta av kol- och väteatomer förenade av varandra mer eller mindre linjära. De har en metylgrupp vid en av sina terminala ändar och en sur karboxylgrupp i den andra, för vilken de kallas "fettsyror".
Sammanfattning av syntesen av fettsyror (källa: Mephist.Org/licenser/BY-SA/3.0) via Wikimedia Commons) Lipider är molekyler som används av olika cellbiosyntetiska system för bildning av andra mer komplexa molekyler såsom:
- membranfosfolipider
- triglycerider för energilagring och
- Förankringarna av några speciella molekyler som finns på ytan av många typer av celler (eukaryoter och prokaryoter)
Dessa föreningar kan existera som linjära molekyler (med alla kolatomer mättade med vätemolekyler), men de i linjär kedja kan också observeras och med vissa mättnader, det vill säga med dubbelbindningar mellan dess kolatomer.
Mättade fettsyror kan också hitta grenade kedjor, vars struktur är lite mer komplex.
De molekylära egenskaperna hos fettsyror är avgörande för deras funktion, eftersom många beror på de fysikalisk -kemiska egenskaperna hos molekylerna som bildas av dessa, särskilt deras smältpunkt, deras grad av förpackningar och deras kapacitet för bicapasbildning.
Således är fettsyrasyntes en extremt reglerad fråga, eftersom det är en serie kritiska sekventiella händelser för cellen från många synpunkter.
[TOC]
Var inträffar syntesen av fettsyror?
I de flesta levande organismer inträffar syntesen av fettsyror i det cytosoliska facket, medan deras nedbrytning huvudsakligen sker mellan cytosol och mitokondrier.
Processen beror på energin som finns i ATP-bindningarna, NADPH-reducerande kraft (vanligtvis härrörande från Penty-fosfatvägen), biotinofaktorn, bikarbonatjoner (HCO3-) och manganjoner.
Hos däggdjursdjur är de viktigaste organen för fettsyrasyntes lever, njurar, hjärna, lungor, bröstkörtlar och fettvävnad.
Det omedelbara syntesunderlaget novo Av fettsyror är acetyl-CoA och slutprodukten är en palmitatmolekyl.
Kan tjäna dig: BHI Agar: Vad är, grund, förberedelse, användningAcetyl-CoA härstammar direkt från bearbetningen av glukolitiska mellanhänder, varför en diet med hög kolhydrater främjar syntesen av lipider (lipogenes) ergo, även av fettsyror.
Enzymer som deltar
Acetyl-CoA är det två-kolsyntesblocket som används för bildning av fettsyror, eftersom flera av dessa molekyler förenas i följd till en malonyl-CoA-molekyl, bildad av karboxylering av en acetyl-CoA.
Det första enzymet i rutten, och ett av de viktigaste med tanke på dess reglering, är den person som ansvarar för karboxylering av acetyl-CoA, känd som acetyl-CoA-karboxylas (ACC), som är en komplex enzymatisk bildas av 4 proteiner och använder biotin som en kofaktor.
Men trots att det finns strukturella skillnader mellan olika arter, är enzymfettsyrasyntaset det som ansvarar för de viktigaste biosyntetiska reaktionerna.
Detta enzym är i verkligheten ett enzymatiskt komplex bestående av monomerer som har de sju olika enzymatiska aktiviteterna, som är nödvändiga för förlängning av fettsyra vid "födelse".
De sju aktiviteterna i detta enzym kan listas enligt följande:
- AV: Acilo -grupptransportprotein
- Acetyl-ACP Transacetilasa (PÅ)
- p-cetoacil-ACP-syntas (Ks)
- Malonyl-CoA-ACP-transferas (MT)
- p-cethoacil-ACP-reduktas (KR)
- p-hydroxyacil-ACP-dehydratas (HD)
- Enoil ACP Redtase (Er)
I vissa organismer såsom bakterier bildas till exempel syntasfettkomplexet av oberoende proteiner som är associerade med varandra, men kodas av olika gener (fettsyrasystem syntas typ II).
 Fettsyrasyntesa av jäst (källa: xiong och., Lomakin, jag.B., Steitz, t.TILL. / Public Domain, via Wikimedia Commons)
Fettsyrasyntesa av jäst (källa: xiong och., Lomakin, jag.B., Steitz, t.TILL. / Public Domain, via Wikimedia Commons) I många eukaryoter och vissa bakterier innehåller emellertid multienzymet flera katalytiska aktiviteter som är separerade i olika funktionella domäner, i en eller flera polypeptider, men som kan kodas med samma gen (fettsyrasyntas typ I).
Stadier och reaktioner
De flesta av de studier som genomförts med avseende på syntesen av fettsyror involverar resultaten som gjorts i bakteriemodellen, men mekanismerna för syntes av eukaryota organismer har också studerats med en del djup.
Det är viktigt att nämna att fettsyra -systemet typ II kännetecknas av att alla fett acylförmedlare kovalent går med i ett litet sized -protein känt som acyltransportproteinet (ACP), som transporterar dem från ett enzym till nästa.
Kan tjäna dig: ABO -system: inkompatibilitet, arv och bevisI eukaryoter, tvärtom, är ACP -aktivitet en del av samma molekyl, och förstå att enzymet själv har en speciell plats för föreningen mellan mellanhänder och deras transport genom de olika katalytiska domänerna.
Unionen mellan proteinet eller AVS -delen och den feta acil.
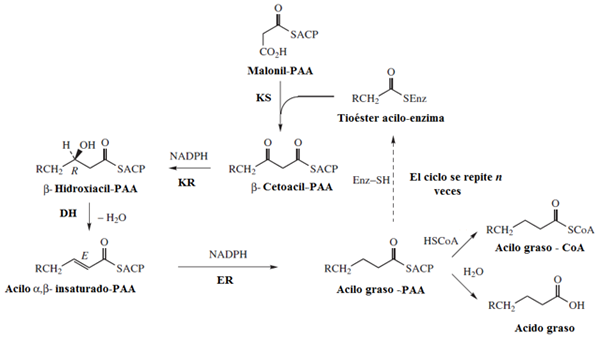
- Ursprungligen är acetyl-CoA-karboxylas (ACC) -enzymet ansvarigt för att katalysera det första steget i ”engagemang” i syntesen av fettsyror som, som nämnts, innebär karboxylering av en acetyl-CoA-molekyl för att bilda mellanhanden av 3 kolatomer som är kända Som malonyl-CoA.
Syntasfettsyrakomplexet får acetyl- och malonilgrupper, som måste fylla "tiol" -platserna korrekt.
Detta sker initialt för överföring av acetyl-CoA till Cisteína SH-gruppen i ß-ZO-beacil-ACP-syntas, reaktion katalyserad av acetyl-CoA-ACP-transacetilas.
Malonylgruppen överförs från malonyl-CoA till SH-gruppen av ACP-proteinet, en händelse medierad av malonyl-CoA-ACP-transferasenzymet och bildar malonyl-ACP.
- Initieringen av förlängningen av fettsyra vid födseln består av kondensation av malonil-ACP med en acetyl-CoA-molekyl, en reaktion regisserad av ett enzym med p-cetoacyl-ACP-aktivitetssyntas. I denna reaktion bildas acetoacetyl-ACP och en CO2-molekyl frigörs.
- Töjningsreaktioner förekommer i cykler där 2 kolatomer tillsätts samtidigt, att varje cykel består av en kondensation, en reduktion, en dehydrering och en andra reduktionshändelse:
- Kondensation: Acetyl- och malonylgrupper kondenseras för att bilda acetoacetyl-ACP
- Reduktion av karbonylgruppen: Den karbonilgruppen av kol 3 i acetoacetyl-ACP reduceras, och bildar D-p-hydroxibuteril-ACP, reaktion katalyserad av p-cetoacyl-ACP-reduktas, som NADPH använder som elektron donor.
- Dehydrering: Hydrogener mellan kol 2 och 3 av den föregående molekylen avlägsnas och bildar en dubbelbindning som slutar med produktionen av The Production of the trans-∆2-butenoil-ACP. Reaktionen katalyseras av p-hydroxyacil-ACP-dehydratas.
- Dubbel länkreduktion: dubbelbindningen av trans-∆2-Butenoil-ACP reduceras till bild av butiril-ACP genom verkan av ilska-ACP-reduktaset, som NADPH också använder som ett reducerande medel.
Det kan tjäna dig: Chiapas flora och fauna: representativa arterFör att fortsätta med förlängningen måste en ny malonylmolekyl gå med i ACP -delen av fettsyrakomplexsyntasen och börjar med kondensationen av detta med butyralgruppen som bildas i den första syntescykeln.
 Palmitato -struktur (källa: edgar181 / allmän domän, via Wikimedia Commons)
Palmitato -struktur (källa: edgar181 / allmän domän, via Wikimedia Commons) I varje töjningssteg används en ny malonyl-CoA-molekyl för att odla kedjan i 2 kolatomer och dessa reaktioner upprepas tills rätt längd har uppnåtts (16 kolatomer), varefter ett enzymliberas-tioesteras full fettsyra genom hydratisering.
Palmitate kan därefter behandlas av olika typer av enzymer som modifierar deras kemiska egenskaper, det vill säga de kan införa omättnad, förlänga deras längd etc.
Reglering
Liksom många biosyntetiska eller nedbrytningsvägar regleras fettsyrasyntes av olika faktorer:
- Det beror på närvaron av bikarbonatjoner (HCO3-), av vitamin B (biotin) och acetyl-CoA (under den första passagen av rutten, vilket innebär karboxylering av en acetyl-CoA-molekyl med hjälp av en karboxiard-mellanhand av biotin för att bilda malonyl-CoA).
- Det är en rutt som inträffar som svar på cellulära energiegenskaper, eftersom när det finns tillräcklig mängd "metaboliskt bränsle" omvandlas överskott till fettsyror som lagras för senare oxidation i ögonblick av energiunderskott.
När det gäller regleringen av acetyl-CoA-karboxylasenzymet, som representerar det begränsande steget för hela vägen, hämmas detta av palmitail-CoA, syntesens huvudprodukt.
Dess toostalaktivator är å andra sidan citrat, som leder metabolism från oxidation till dess syntes för lagring.
När acetyl-CoA- och ATP-mitokondriella koncentrationer ökar, transporteras citrat till cytosol, där det är så föregångare för cytosolisk syntes av acetyl-CoA och en alkalistisk aktiveringssignal för acetyl-CoA-karboxylas.
Detta enzym kan också regleras av fosforylering, händelse som skjuts av den hormonella verkan av glukagon och epinefrin.
Referenser
- Mcgenity, t., Van der Meer, J. R., & De lorenzo, v. (2010). Handbok för kolväte och lipidmikrobiologi (P. 4716). K. N. Timmis (Ed.). Berlin: Springer.
- Murray, r. K., Granner, D. K., Mayes, s. TILL., & Rodwell, V. W. (2014). Harpers illustrerade biokemi. McGraw-hill.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger Principles of Biochemistry (pp. 71-85). New York: Wh Freeman.
- Numa, s. (1984). Fettsyrametabolism och dess reglering. Annars.
- Rawn, j. D. (1989). Biokemi-internationell upplaga. North Carolina: Neil Patterson Publishers, 5.
- « Systematiskt fel hur man beräknar det, i kemi, i fysik, exempel
- Omättade fettsyroregenskaper, struktur, funktioner, exempel »