

Steroidhormoner struktur, syntes, verkningsmekanism
- 1764
- 377
- Johan Johansson
De Steroidhormoner De är ämnen gjorda av inre utsöndringskörtlar och som dumpar direkt i cirkulationslordrenten, vilket leder dem till vävnaderna där de utövar sina fysiologiska effekter. Dess generiska namn härstammar från det faktum att ha en steroidkärna i dess grundläggande struktur.
Cholesterol is the precursor substance from which all steroid hormones are synthesized, which are grouped into progestaging (for example, progesterone), estrogens (stronation), androgens (testosterone), glucocorticoids (cortisol), mineralocorticoids (aldosterone) and vitamin D.
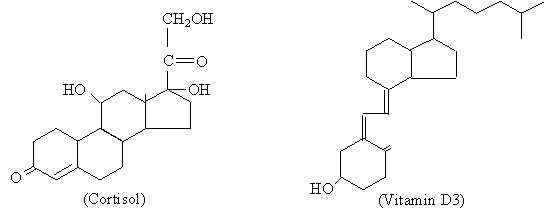
Jämförelse av strukturen hos ett steroidhormon (kortisol) med en molekyl av samma kemiska natur (vitamin D3) (Källa: Den ursprungliga uppladdaren var Palladius vid engelska Wikipedia. [Public domain] via Wikimedia Commons) Även om de olika steroidhormonerna som finns bland dem molekylära skillnader som är de som ger deras olika funktionella egenskaper, kan det sägas att de har en grundstruktur som är gemensam för dem och som representeras av cyklopentanoperhydrofenantreno av 17 kolatomer.
[TOC]
Steroidstruktur
Steroider är organiska föreningar av mycket mångfaldig natur som har gemensamt vad som kan betraktas som en stamkärna som består av sammansmältningen av tre ringar av sex kolatomer (cyklohexan) och en av fem kolatomer (cyklopentano).
Denna struktur är också känd som "Cyclopentanoperhydrofenantreno". Eftersom ringarna är förenade ömsesidigt är de totala kolatomerna som utgör det 17; Men mest naturlig steroid.
 Schema för de fyra ringarna i Cyclopentanoperhidrofenantreno Cykropisk struktur (källa: Neurotokeker [Public Domain] via Wikimedia Commons)
Schema för de fyra ringarna i Cyclopentanoperhidrofenantreno Cykropisk struktur (källa: Neurotokeker [Public Domain] via Wikimedia Commons) Många av de naturliga steroidföreningarna har också en eller flera grupper med alkoholhaltig funktion och får därför namnet på steroler. Bland dem är kolesterol, som har en alkoholfunktion i kol 3 och en lateral kolvätekedja med 8 kol -fästade kolatomer 17; atomer som är numrerade från 20 till 27.
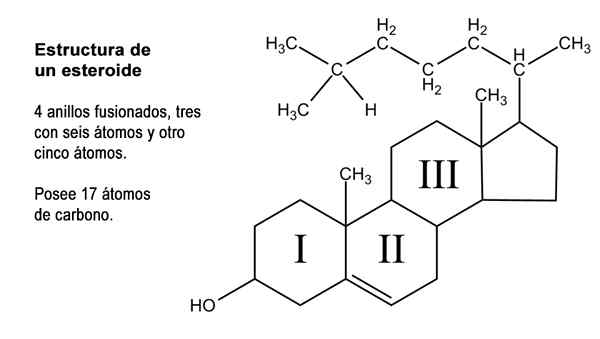 Steroidstruktur. Modifierad bild av Marcotolo/CC BY-S (https: // CreativeCommons.Org/licenser/BY-SA/2.5)
Steroidstruktur. Modifierad bild av Marcotolo/CC BY-S (https: // CreativeCommons.Org/licenser/BY-SA/2.5) Förutom dessa 17 kol kan steroidhormoner ha i sin struktur 1, 2 eller 4 fler av dessa atomer, så tre typer av steroider erkänns, nämligen: C21, C19 och C18.
C21
C21, såsom progesteron och binjurekortikosteroider (glukokortikoid och mineralokortikoider) härrör från "gravan". Detta har 21 kolatomer eftersom vid 17 av basringen läggs de två av metylgrupperna av kol 13 och 10, och två kolhydrater i sidokedjan fäst vid C17 som ursprungligen, i kolesterolet, var det 8 kolhydrater.
C19
C19 motsvarar könshormoner med androgen aktivitet och härstammar från "androstano" (19 kolatomer), som är strukturen som återstår när gravan förlorar de två kolhydrater i sidokedjan i C17, som ersätts av en hydroxyl eller hydroxyl en ketonisk grupp.
C18
C18 -steroiderna är kvinnliga eller östrogenhormoner som huvudsakligen syntetiseras i kvinnliga gonader och vars framträdande egenskap, med avseende på de andra två typerna av steroider, är frånvaron av metyl som finns i det senare bundet till kol i position 10.
Under syntesen från kolesterol produceras enzymatiska modifieringar som förändrar antalet kol och främjar dehydrogenering och hydroxilationer av specifika kolhydrater i strukturen.
Syntes
Steroidhormonproducerande celler är huvudsakligen belägna i barken av binjurarna, där glukokortikoider såsom kortisol, mineralokortikoider såsom aldosteron och manliga könshormoner såsom dehydroepiandrosteron och androstenaeda.
Manliga sexuella gonader ansvarar för produktionen av androgener som inkluderar de redan nämnda hormonerna och testosteron, medan äggstocksäckar som når mognad producerar progesteron och östrogen.
Kan tjäna dig: hälften av mig: vad är, grund, förberedelse, användningSyntesen av alla steroidhormoner börjar från kolesterol. Denna molekyl kan syntetiseras av steroidhormoner som producerar celler, men mest erhålls dessa celler från lipoproteiner med låg densitet (LDL) närvarande i den cirkulerande plasma.
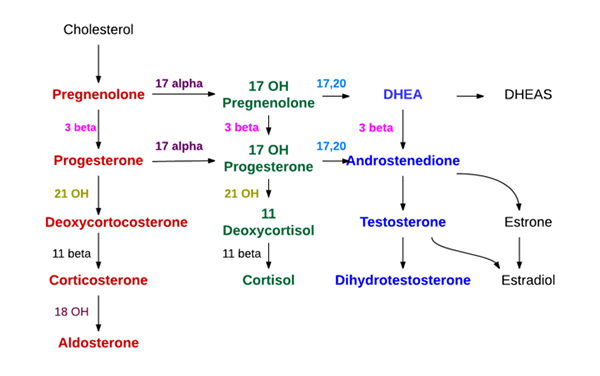 Syntes av binjurhormoner (källa: endokrin läkare [CC BY-SA (https: // Creativecommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons)
Syntes av binjurhormoner (källa: endokrin läkare [CC BY-SA (https: // Creativecommons.Org/licenser/BY-SA/4.0)] via Wikimedia Commons) - Syntes på binjurens cortexnivå
Tre lager kännetecknas i binjurbarken, känd från utsidan som glomerulära, fasculära och retikulära områden, respektive.
I glomerulära är de grundläggande syntetiserade mineralokortikoider (aldosteron), i de fascikulära glukokortikoiderna såsom kortikosteron och kortisol, och i retikulära androgener såsom dehydroepiandrosteron och androsten.
Glukokortikoidsyntes
Det första steget i syntesen inträffar i mitokondrierna och består i verkan av ett enzym som kallas kolesteroldemollas, som tillhör superfamiljen av cytokrom P450 och även känd som "P450SCC" eller "CYP11A1", vilket främjar eliminering av 6 av 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av de 6 av 6 av de 6 av 6 av 6 av 6 av 6 av 6 av 6 av 6 av 6 av 6 av 6 av 6 Kolatomer i sidokedjan kopplad till C17.
Med verkan av reklöst blir kolesterol (27 kolatomer) en gravidolon, som är en förening med 21 kolatomer och representerar den första av steroiderna av typ C21.
Den gravneenolonen rör sig till det släta endoplasmiska retikulum, där genom verkan av enzymet 3p-hydroxiesteroiddehydrogenas.
Genom verkan av 21p-hydroxylaset, även kallad “P450C21” eller “CYP21A2”, är progesteronet hydroxyle i kol 21 och förvandlas till 11-dexikortikosteron, som återgår till mitokondrierna och till vilket enzymet 11p-hydroxylas ("" " P450C11 ”eller“ CYP11B1 ”) konverterar till kortikosteron.
En annan synteslinje i fascikulärzonen och som inte slutar i kortikosteron, men i kortisol, inträffar när gravanolonen eller progesteronet hydroxyleras i position 17 av 17a-hydroxylaset ("P450C17" eller "CYP17") och omvandlas till 17-hydroxenolon eller 17-hydroxiprgesteron.
Samma enzym som redan nämnts, dehydrogenas av 3p-hydroxiesteroider, som omvandlar gravenolon till progesteron, konverterar också 17-hydroxipnenolon i 17-hydroxiprgesteron.
Det senare bärs successivt av de två sista enzymerna på vägen som producerar kortikosteron (21p-hydroxylas och 11p-hydroxylas) till deoxyxikortisol respektive kortisol.
Glukokortikoidåtgärder
De viktigaste glukokortikoiderna som produceras i fascikulär zonen i binjurcortex är kortikosteron och kortisol. Båda ämnena, men särskilt kortisol, visar ett brett spektrum av handlingar som påverkar metabolism, blod, försvar och sårläkningssvar, benmineralisering, matsmältningskanal, cirkulationssystem och lungor.
När det gäller metabolism stimulerar kortisol lipolys och frisättning av fettsyror som kan användas i levern för bildning av ketonkroppar med låg densitet (LDL); minskar glukosuppsamlingen och lipogenesen i fettvävnad och insamling och användning av glukos i muskeln.
Det främjar också proteinkatabolism i periferin: i kontaktvävnaden, muskeln och benmatrisen, som frigörs aminosyror som kan användas i levern för syntes av plasmaproteiner och för glukoneogenes. Stimulerar dessutom tarmsglukosabsorption genom att öka produktionen av SGLT1 -transportörer.
Accelerera glukos tarmsabsorption, förstärkt leverproduktion och minskad användning av detta kolhydrat i muskel- och fettvävnad gynnar en höjd av plasmaglukosnivåer.
När det gäller blodet gynnar kortisol koaguleringsprocessen, stimulerar bildningen av neutrofiler och hämmar den hos eosinofiler, basofiler, monocyter och lymfocyter T. Det hämmar också befrielsen av inflammationsförmedlare som prostaglandiner, interleukiner, lymfociner, histamin och serotonin.
Kan tjäna dig: kemiorreceptorI allmänhet kan det sägas att glukokortikoider stör immunsvaret, så att de kan användas terapeutiskt i de fall där detta svar är överdrivet eller olämpligt, som i fallet med autoimmuna sjukdomar eller i organtransplantationer för att minska avslag.
- Androsyntes
Androgen syntes vid binjurens cortex-nivå är främst på nivån i retikulär zon och från 17-hydroxipregneolon och 17-hydroxiprgesteron.
Samma enzym 17a-hydroxylas, som producerar de två nyligen nämnda ämnena, har också 17,20 LIASA-aktivitet, som undertrycker de två kol i sidokedjan i C17 och ersätter dem med en keto-grupp (= o).
Denna sista åtgärd minskar antalet kol och producerar steroider av typ C19. Om handlingen inträffar på 17-hydroxnenolonen är resultatet dehydroepiandrosteron; Om det tvärtom, det drabbade ämnet är hydroxiprgesteron, kommer produkten att vara androstenen.
Båda föreningarna är en del av de så kallade 17-zosteroiderna, eftersom de har en ketongrupp i kol 17.
3p-hydroxisteroid dehygenas omvandlar också dehydroepiandrosteron i androsten.
Mineralocorticoid -syntes (Aldosteron)
Den glomerulära zonen saknar enzymet 17a-hydroxylas och kan inte syntetisera de 17-hydroxiesteroida föregångarna till kortisol och sexhormoner. Det har inte heller 11p-hydroxylas, men ett enzym som kallas aldosteronsyntes som kan producera sekventiellt kortikosteron, 18-hydroxikorik och aldosteronmineralokortikoid.
Mineralokortikoidåtgärder
Den viktigaste mineralokortikoiden är det syntetiserade aldosteronet i det glomerulära området i binjurbarken, men glukokortikoider visar också mineralokortikoidaktivitet.
Aldosteron -mineralokortikoidaktiviteten utvecklas på nivån för det rörformiga epitelet i de distala nefronkroppsvätskorna.
- Syntes av manliga könssteroider i testiklarna
Androgentestikulär syntes sker på Leydig -cellernivån. Testosteron är det huvudsakliga androgena hormonet som produceras i testiklarna. Dess syntes innebär den initiala produktionen av androstenediona som tidigare beskrivits för syntesen av androgener på nivån av binjurebarken.
Androsteneciona omvandlas till testosteron genom verkan av enzymet 17p-hydroxiesteroid dehydrogenas, som ersätter ketongruppen av kol 17 med en hydroxylgrupp (OH).
I vissa vävnader som fungerar som ett mål för testosteron reduceras detta med ett 5a-reduktas till dihydrotestosteron, med större androgen kraft.
- Syntes av kvinnliga könssteroider i äggstockarna
Denna syntes sker cykliskt åtföljande de förändringar som inträffar under den kvinnliga sexuella cykeln. Syntesen sker i follikeln att under varje mogen cykel för att frigöra ett ägg och sedan producera motsvarande luteumkropp.
Östrogener syntetiseras i de korniga cellerna i den mogna follikeln. Den mogna follikeln har celler i teak som producerar androgener såsom androstenedion och testosteron.
Dessa hormoner sprids till de angränsande granulösa cellerna, som har aromatasenzymet som gör dem till strikt (E1) och 17p-stradiol (E2). Från båda syntetiseras Striol.
Åtgärder av könssteroider
Andogener och östrogener har som huvudfunktion utvecklingen av manliga respektive kvinnliga sexuella karaktärer. Andogener har anabola effekter som främjar syntesen av strukturella proteiner, medan östrogener gynnar ossifieringsprocessen.
Östrogener och progesteron som släpps under den kvinnliga sexuella cykeln riktas.
Kan tjäna dig: Flora och fauna från Baja California SurHandlingsmekanism
Om du behöver uppdatera minnet om mekanismen för hormonverkan rekommenderas att du visualisera följande video innan du fortsätter.
Verkningsmekanismen för steroidhormoner är ganska lik i dem alla. När det gäller lipofila föreningar upplöses de utan svårigheter i lipidmembranet och penetrerar cytoplasma i deras vita celler, som har specifika cytoplasmiska receptorer för det hormon som de måste svara.
När hormon-mottagskomplexet bildas korsar det kärnmembranet och förenas i genomet, på sätt som en transkriptionsfaktor, med ett element av svar på hormonet (HRE) eller primär svarsgen, som du kan reglera reglering Andra sekundära svargener.
Det slutliga resultatet är att främja transkript och syntes av RNA -budbärare som översätts till ribosomerna i den grova endoplasmiska retikulum som slutar syntetisera de hormoninducerade proteinerna.
Aldosteron som ett exempel
 Aldosteronmolekyl
Aldosteronmolekyl Aldosteronets verkan utövas huvudsakligen på nivån på den sista delen av det distala röret och i uppsamlingskanalerna, där hormonet främjar reabsorption av Na+ och utsöndring av k+.
I det luminalmembranet i de viktigaste rörformiga cellerna i denna region finns epitelkanaler av Na+ och K+ -kanaler av den engelska typen (från engelska Yttre medullär kaliumkanal).
Det basolaterala membranet har Na+/K+ ATPASA -pumpar som kontinuerligt tar Na+ från cellen till det basolaterala interstitiella utrymmet och introducerar K+ i cellens inre. Denna aktivitet håller den intracellulära koncentrationen av Na+ mycket låg och gynnar skapandet av en koncentrationsgradient för denna jon mellan tubulens ljus och cellen.
Denna lutning tillåter Na+ att röra sig mot cellen genom epitelkanalen, och när Na+ passerar ensam, för varje jon som rör sig finns det en okompenserad negativ belastning som får tubulusens ljus att bli negativt med avseende på interstitium. Det vill säga en transepitelial potentialskillnad skapas med det negativa ljuset.
Denna negativitet hos ljuset gynnar utgången av K+ som rörde sig genom dess större koncentration i cellen och ljusets negativitet utsöndras mot tubulens ljus för att äntligen utsöndras. Det är denna Na+ resorptionsaktivitet och utsöndring av K+ som regleras av aldosteronets verkan.
Aldosteronet som finns i blod och frisatt från den glomerulära zonen som svar på verkan av angiotensin II, eller till hyperkalemi, tränger in i huvudcellerna och sammanfogar dess intrakitoplasmiska receptor.
Detta komplex når kärnan och främjar transkriptionen av generna vars uttryck kommer att hamna med att öka syntesen och aktiviteten för Na+/K+-pumpar, av Na+-epitelkanalerna och ROMK -kanalerna för K+, såväl som andra proteiner mer. Svar som kommer att ha kvarhållandet av Na+ i organismen och ökningen av urinutsöndring av k+.
Referenser
- Ganong WF: Adrenal Medulla & Adrenal Cortex, 25: e upplagan. New York, McGraw-Hill Education, 2016.
- Guyton AC, Hall JE: Adrenokortikala hormoner, i Lärobok för medicinsk fysiologi , 13: e upplagan, AC Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Lang F, Verrey F: Hormon, i Physiologie des Menschen Mite Pathophysiologie, 31 ED, RF Schmidt et al (eds). Heidelberg, Springer Medizin Verlag, 2010.
- Voigt K: Endokrines System, i: Physiologie, 6: e upplagan; R KLINKE et al (eds). Stuttgart, Georg Thieme Verlag, 2010.
- Widmaier EP, Raph H och Strang KT: Kvinnlig reproduktiv fysiologi, i Vanders mänskliga fysiologi: Mekanismerna för kroppsfunktion, 13: e upplagan; EP Widmaier et al (eds). New York, McGraw-Hill, 2014.
- « Statisk friktionskoefficient, exempel, träning
- Ampère formel och ekvationer lag, demonstration, övningar »